Research Article
The Mechanism of Bacterial Translocation after Major Hepatectomy in Cirrhotic Rats
Jian Liang1,2, Zhi-qiang Ye3, Shu-xian Chen1, Chu-si Wan1 and Wei-dong Pan1*
1Department of Hepatopancreatobiliary Surgery, The Third Affiliated Hospital of Sun Yat-sen University, China
2Department of General Surgery, Hospital of Traditional Chinese Medicine of Zhongshan, China
3Department of Emergency, The Third Affiliated Hospital of Sun Yat-sen University, China
*Corresponding author: Wei-dong Pan, Department of Hepatopancreatobiliary Surgery, The Third Affiliated Hospital of Sun Yatsen University, China
Published: 28 Jun, 2018
Cite this article as: Liang J, Ye Z-q, Chen S-x, Wan C-s,
Pan W-d. The Mechanism of Bacterial
Translocation after Major Hepatectomy
in Cirrhotic Rats. Clin Surg. 2018; 3:
2002.
Abstract
This study was designed to investigate the mechanism underlying bacterial translocation after major
hepatectomy in cirrhotic rats. Sixty experimental cirrhotic rats were randomly divided into SO, PH
and PHP groups. Portal pressure, the composition of the intestinal micro flora, the levels of D-lactic
acid, endo toxins, inflammatory factors, and tight junction proteins were analyzed in the three
groups of animal. Portal pressure of the PH rats was significantly higher (22.48 ± 0.882 cm H2O)
than that of the SO (14.3 ± 1.25 cm H2O) and PHP rats (16.6 ± 1.02 cm H2O) (P<0.05). The PH
rats also showed significantly higher levels of serum LPS and D-lactic acid (52.56 ± 2.08 EU/ml and
60.87 ± 4.44 mg/L) than the SO (48.28 ± 1.45 EU/ml and 37.23 ± 1.71 mg/L) and PHP rats (47.84 ±
3.45 EU/ml and 40.83 ± 1.82 mg/L) (P<0.05). ZO-1 and Claudin-1 levels were significantly higher in
the SO and PHP rats than in the PH rats (P<0.05), and the expression levels of TNF-α, IFN-γ, and
MLCK were significantly lower in the SO and PHP rats than in the PH rats (P<0.05). Portal pressure
significantly increased after major hepatectomy in cirrhotic rats, leading to intestinal stagnant
anoxia, the accumulation of local metabolic products and inflammatory factors, the induction of
intestinal epithelial cell apoptosis, and changes in the composition of the gut flora. These effects
collectively resulted in tight junction functional disorders.
Keywords: Cirrhosis; Major hepatectomy; Bacterial translocation; Gut barrier
Introduction
Postoperative infection is a common complication of major hepatectomy, after which it represents an important cause of patient deaths [1]. Studies have shown that postoperative infection following major hepatectomy is closely associated with intestinal Bacterial Translocation (BT) [2,3] and leads to increased mortality. The reasons that these endogenous infections occur remain elusive, and no single determinant has been recognized. Studies have shown that portal venous pressure sharply increases during major hepatectomy and that portal hypertension can lead to congestion and edema of the bowel intestinal barrier. These changes may enable the translocation of intestinal tract bacteria from the intestinal lumen to tissues and organs in the abdominal cavity, where they can stimulate a series of inflammatory responses [3]. Determining the mechanism underlying BT and identifying ways to maintain the integrity of the intestinal barrier are therefore important goals for research on major hepatectomy. In the present study, we investigated the changes that occurred in the mechanical and bacterial barriers of the intestines of cirrhotic rats that underwent major hepatectomy with the goal of identifying the mechanisms underlying bacterial translocation.
Materials and Methods
Experimental animals
In the present study, the Specific Pathogen-Free (SPF) levels of 80 male Sprague Dawley (SD)
rats weighing 80 g to 100 g were measured. The rats were purchased from the animal experimental
center of Sun Yat-sen University and fed at the animal experimental center of Sun Yat-sen
University, North Campus, in a SPF-level barrier environment (experimental animal usage license
No. SYXK (Guangdong 2012-0081). The rats were fed under artificial lighting every 12 hr. Housing
was maintained at 24°C to 26°C and 50% to 60% relative humidity. After the experiments, the rats
were disposed of in accordance with the National Institutes of Health Guidelines for the Care and
Use of Laboratory Animals. All protocols used in this study were approved by the Sun Yat-sen
University Animal Care and Use Committee.
Preparation of animal model
A 40% liquid CCl4 soy bean oil mixture (0.4 ml/100 g body
weight) was used to perform abdominal subcutaneous multi-point
injections in the rats twice a week. After 10 weeks, 80% of the rats
were successfully induced to develop cirrhosis.
Animal groups
A total of 60 cirrhotic rats were randomly divided into Sham
Operation (SO), major Hepatectomy (PH) and major Hepatectomy
Plus Propranolol (PHP) groups. Each group consisted of 20 rats. A
total of 10 rats in each group were selected and measured for portal
pressure. The remaining 10 rats were used for detection.
Operation methods and collected materials
The rats were subjected to preoperative fasting for 8 h and provided
drinking water ad libitum. Ether was administered intranasally to
induce anesthesia, and iodine was used to disinfect the skin in the
operation field. A median abdominal incision was performed in the
abdomen to expose the liver. In the SO group, the ligament around
the liver was cut, and the abdomen was then closed. In the PH group,
a modified major hepatectomy was performed on the rats according
to the protocol described by Kamada and Andersson [4,5], in which
the left inner and outer lobes and the middle lobe were cut from the
liver, resulting in a resection of approximately two-thirds of the liver.
In the PHP group, a major hepatectomy was conducted, and the rats
were then perused with 2 ml of a 5 mg/ml propranolol saline solution
at 12 hr and 24 hr after the surgery.
Portal pressure determination
During laparotomy, a small catheter was inserted directly into
the portal vein, and portal pressure was measured through a pressure
transducer that was connected to a biological signal acquisition
system and registered using a multi channel recorder (BL-420F,
Chengdu Technology and Market Co., Ltd. Chengdu, China). The
results are expressed as cmH2O.
Materials collected for detection
At 24 h post-operation, the abdomen was re-opened and materials
were collected under sterile conditions.
Plasma D-Lactate levels and endotoxin levels
A total of 2 ml of blood was drawn from the inferior vena cava.
Plasma was obtained via separation in a centrifuge. The samples were
tested with ELISA assays according to the manufacturer’s instructions
(Shanghai, China, Bio-Techne China Co. Ltd.). D-lactate levels are
expressed in mg/L, and endotoxin levels are expressed as EU/ml.
Fecal bacteria detection by PCR
Fecal DNA extraction
Approximately 0.5 g of fecal matter was collected and weighed.
A 500 μL volume of protease was added to the samples during lysis,
and the solutions were mixed and digested overnight. Tris-balanced
phenol chloroform and absolute ethyl alcohol were then used to
extract DNA. The program Primer 5.0 was used to design appropriate
bacteria genus PCR primers, and the specificity of the primer
sequences was compared to the corresponding bacterial sequences in
GenBank.
PCR primer design
Enterococcus: F: GGTTTCCGCCCTTCAGTG, R:
GAAGCTCTATCTCTAGAGTGGTCA
Clostridium: F: TAATGACGGTACTTTGGAGGA, R:
CGTAGCCCTTTCTACCTGC
Bacteroides: F: GCAGCATATTTGTAGCAATACAGAT, R:
CTATACCATCGGGTATTAATCTTTC
Bifidobacteria: F: GAGCAAGCCTTCGGGTGAGT, R:
GGCCCCACATCCAGCGTC
Colon Bacillus: F: ACGGTAACAGGAAGCAGCTT, R:
CCCTCTTTGGTCTTGCGA
Klebsiella pneumonia: F: GATGAAACGACCTGATTGCATTC,
R: CCGGGCTGTCGGGATAAG
23S reference: F: AGCGAAAGACAGGTGAGAATCC, R:
CCTATCGGCCTCGGCTTAG
Fecal bacteria PCR analysis
The amplification system consisted of 0.2 μL of each forward and
reverse primer (10 μM), 12.5 μL of Premix Ex TaqTM (2X), 3 μL of
template DNA, and enough ddH2O to reach a final reaction volume
of 25 μL. The following PCR conditions were used: one cycle of
initial denaturation at 95°C for 15 min followed by 40 cycles of PCR
amplification at 94°C for 30 s, 60°C for 30 s, and 72°C for 90 s.
Detection of bacteria in mesenteric lymph nodes
Several Mesenteric Lymph Node (MLN) samples were obtained,
and the protocol used to detect bacteria was the same that was used
to detect fecal bacteria. The following bacterial primer sequences were
used:
F: CTCCTACGGGAGGCAGCAGT and R:
ATTACCGCGGCTGCTGGCAC
Small intestine and liver HE staining
The resected distal ileum segments were fixed in neutral-buffered
formalin and then processed for histological analysis. The slides were
stained with Hematoxylin and Eosin (HE) and analyzed under an
optical microscope (Leica DMI4000B) by a single pathologist who
was blinded to the sample identities. The pathological changes that
occurred in the small intestine mucosa were evaluated using Chiu’s
scoring method [6]: 0 points, normal; 1 point, the subcutaneous space
on the villus tip was widened; 2 points, the subcutaneous space on the
villus tip was further expanded, the villus tip epithelium was elevated,
and the intrinsic membrane was peeling; 3 points, the villus epithelium
was shedding in bulk; 4 points, the epithelium was completely shed,
leaving only an inherent film; and 5 points, the inherent film was
cracked, and bleeding and ulceration were observed.
Western blot detection of ZO-1 and Claudin-1 in small
intestine tissues
Resected distal ileum segments were stored at -80°C. A sample
(100 μg) of intestinal mucosal tissue was collected and added to a
solution containing lysate and buffer. The mixture was homogenized
and centrifuged. The supernatant was collected, and 10 μL of the
diluted samples and 5 μL of a prestained marker were loaded onto
gels. After denaturation, electrophoresis, film transfer and closure
had been performed, the corresponding primary and secondary
antibodies were added. GAPDH was used as the inner reference.
After the samples were incubated and developed, a quantitative
analysis was conducted.
Immunohistochemical detection of TNF-α, IFN-γ and
MLCK in small intestine tissues
The distal ileum tissue was collected, embedded, and fixed.
Next, the samples were subjected to antigen repair, endogenous
peroxidase activity was blocked, and the tissue was incubated with
the corresponding primary and secondary antibodies. Subsequently,
SABC reagent was added, and the tissues were incubated, colored
with DAB, and restrained. In the negative control, the antibody was
replaced with PBS. The results of the imunohistochemical analysis
were interpreted in semi-quantitative mode using a double-blind
method, and cells that appeared as red granules were considered to
be positively labeled. Over 75% of the cells that were stained were
strongly positive (+++), 50% to 75% were positive (++), 10% to 49%
were weakly positive (+), and fewer than 10% were negative.
Statistical analysis
Portal pressure, endotoxin concentration, and D-lactate, ZO-
1and Claudin-1 level are expressed as the mean ± standard error,
and differences between two groups were analyzed using t-tests
for two independent samples. The results of PCR to detect fecal
components and MLN bacteria were analyzed using a gene/inner
reference gene method for the expression level analysis. We used
Chiu’s scoring system for small intestine tissue samples to compare
the two groups of independent samples (i.e., the Wilcoxon test). The
results of immunohistochemistry in the ileum for TNF-α, IFN-γ and
MLCK showed that they were expressed at a strong positive rate,
and Pearson’s χ2 test was used to analyze the detection data. P<0.05
indicated a significant difference.
Figure 1
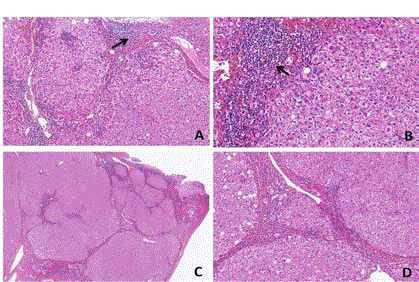
Figure 1
Changes in liver histology in cirrhotic rats. A: Liver cells exhibiting
bridging necrosis in the plates of hepatocytes grown within hepatic lobules
(HE 10×10). B: Liver cells exhibiting piecemeal necrosis in the plates of
hepatocytes grown within hepatic lobules (HE 10×20). C: Normal structures
within hepatic lobules were destroyed, and false lobules had formed (HE
2×10). D: Normal structures within hepatic lobules were destroyed, and false
lobules had formed (high-power image of an area shown in C, HE 10×10).
Figure 2
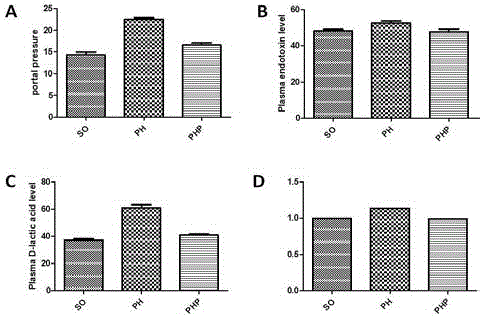
Figure 2
Comparisons of several parameters among three groups. A: A
comparison of portal pressure among the three groups. Portal pressure was
significantly higher in the PH group than in the SO and PHP groups (SO vs.
PH, P<0.01; PHP vs. PH, P<0.01). B: A comparison of plasma endotoxin
levels among the three groups. Plasma endotoxin levels were significantly
higher in the rats in the PH group than in those in the SO and PHP groups
(P<0.05). C: A comparison of plasma D-lactic acid levels among the three
groups. Plasma D-lactic acid levels were significantly higher in the rats in the
PH group than in the SO and PHP groups (P<0.05). D: The total amounts of
bacteria in the mesenteric lymph nodes of the three groups. The total quantity
of bacteria was 14% higher in the MLNs of the PH group than in those in the
SO and PHP groups (P<0.05). There was no significant difference between
the PHP and SO groups.
Figure 3
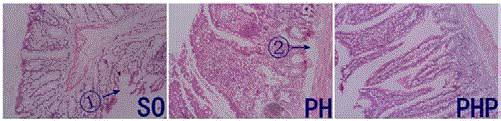
Figure 3
The structure of the intestinal mucosa in the three groups (HE
staining). Arrow ① shows that the mucosal epithelium exhibited reduced
continuity and shortened intestinal epithelial villi. Arrow ② shows a cracked
lamina propria and shrunken intestinal villi. The images were captured at 40×
magnification.
Figure 4
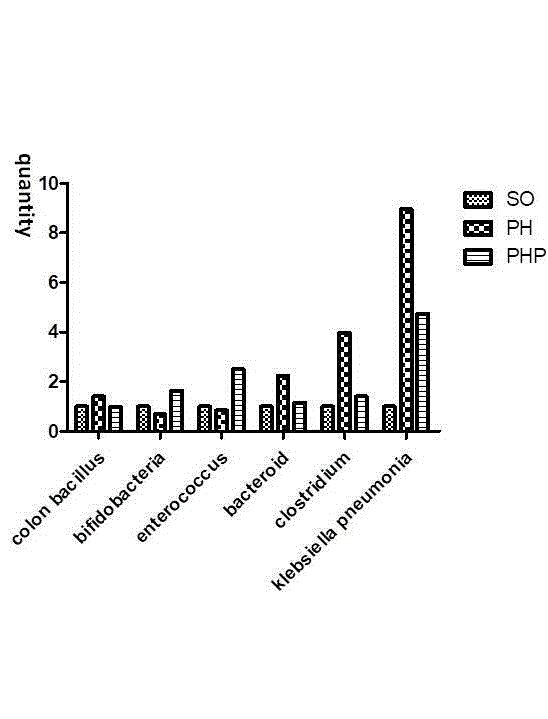
Figure 4
PCR analysis of the composition of fecal bacteria. There were
fewer probiotics, such as Bifidobacteria, in the PH group, whereas there
were more pathogens, such as colon Bacillus, Bacteroides, Clostridium, and
Klebsiella pneumoniae.
Results
Histological changes in the liver
In the 6th week, liver cells exhibited piecemeal (Figure 1A) and
bridging necrosis (Figure 1B) in plates of hepatocytes grown within
hepatic lobules. After 10 weeks, the liver cells were arranged in a
disorderly fashion. The normal structures of the hepatic lobules were
destroyed, and false lobules had formed (Figure 1C, 1D). Slight bile
duct proliferation and extensive fibrous tissue proliferation were also
observed.
Portal pressure
Portal pressure was 14.3 ± 1.25 cmH2O in the SO group. It was
significantly higher in the PH group (22.48 ± 0.882 cm H2O, P<0.01)
than in the SO group and significantly lower in the PHP group (16.6
± 1.02 cmH2O, P<0.01) than in the PH group (Figure 2A).
Plasma endotoxin and D-lactic acid levels
The plasma endotoxin and D-lactic acid levels in the SO group
were 48.28 ± 1.45 EU/ml and 37.23 ± 1.71 mg/L, respectively. In the
PH group, the levels were higher, at 52.56 ± 2.08 EU/ml (SO vs. PH,
P<0.05) and 60.87 ± 4.44 mg/L (SO vs. PH, P<0.05), respectively.
However, plasma endotoxin and plasma D-lactic acid levels were
lower in the PHP group than in the PH group (47.84 ± 3.45 vs. 52.56
± 2.08EU/ml and 40.83 ± 1.82 vs. 60.87 ± 4.44 mg/L, respectively;
P<0.01 for both), and this difference was associated with a lower
prevalence of BT (Figure 2B, 2C).
PCR detection of MLN bacteria
The total amount of bacteria in the MLNs was 14% higher in the
PH group than in the SO and PHP groups (P<0.05), although there
was no significant difference between the PHP and SO groups (Figure
2D).
Small intestine pathology according to the Chiu score
The structure of the intestinal mucosa was generally normal in
the SO group, although there was less continuity in the mucosal
epithelium and shortened intestinal epithelial villi. Above the
villus, the subcutaneous space had widened, the intestinal villus tip
epithelium was elevated, and the fixed film had peeled. The continuity
of the intestinal mucosa was relatively poor in the PH group, in which
the intestinal mucosa epithelium was shedding in bulk and in some
places even completely falling off. The fixed layer was cracked, and its
intestinal villus was smaller than that observed in the SO group. The
Chiu scores showed that the average score for rat mucosal damage
was significantly higher in the PH group (22.55) than in the SO group
(7.35). The score for the structure of the intestinal mucosa in the PHP
group was 16.6, which was between the scores for the PH and SO
groups described above (PHP vs. PH, P<0.05) (Figure 3).
PCR analysis of fecal bacteria
The amounts of fecal Clostridium and Bacteroides and colon
Bacillus species were 125%, 297% and 796% higher, respectively,
while the amounts of Bifidobacteria and Enterococcus species were
31% and 15% lower, respectively, in the PH group than in the fecal
samples obtained in the SO group. However, the amounts of fecal
Clostridium, Bacteroides and colon Bacillus were significantly
lower and the amounts of Bifidobacterium and Enterococcus were
significantly higher in the PHP group (P<0.05) (Figure 4).
Western blot assay
Western blot assays showed that the expression levels of the ZO-1
and Claudin-1 proteins were significantly lower in the PH group
than in the SO and PHP groups, although there were no significant
differences in these levels between the SO and PHP groups (Figure 5).
Immunohistochemical study of the intestinal mucosa
When TNF-α, IFN-γ and MLCK were detected in small intestine
tissues, cells with the appearance of red granules were observed. The
proportions of positive cells were different among three groups. The
proportions of cells that strongly expressed TNF-α, IFN-γ and MLCK
were significantly higher in the PH group (40%, 50%, 70%) than in
the SO (20%, 10%, 20%) and PHP (30%, 20%, 40%) groups (P<0.05;
Figure 6).
Figure 5

Figure 5
Western blots used to detect the ZO-1 and Claudin-1 proteins.
Western blot assays showed that the expression levels of the ZO-1 and
Claudin-1 proteins was significantly lower in the PH group than in the SO
and PHP groups. There was no significant difference between the SO and
PHP groups.
Figure 6
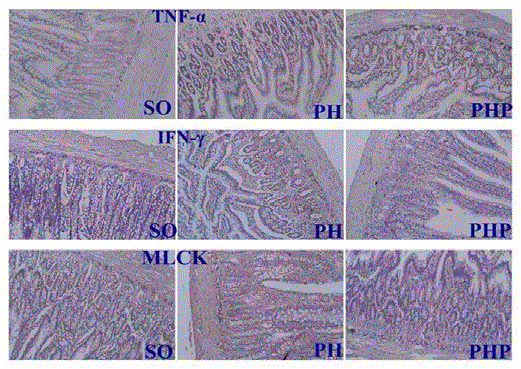
Figure 6
Immunohistochemical study of the intestinal mucosa. Cells
showing positive expression (red granules) were observed. The proportions
of cells that were strongly positive for TNF-α, IFN-γ and MLCK expression
were significantly higher in the PH group than in the SO and PHP groups.
Discussion
Intestinal bacterial translocation is one of the causes of
postoperative infection after major hepatectomy [7]. Many underlying
factors affect how much translocation occurs, including the
reticuloendothelial system [8], which reduces damage to the intestinal
mechanical, bacterial and immune barriers [9-11], increases in the
levels of intestinal tissue inflammatory factors [12], and intestinal
ischemia reperfusion injury [13]. The mechanical barriers presents
the most important barrier in the intestines, and its structure is closely
associated with the expression of members of the ZO and Claudin
families of proteins [14,15]. In the present study, Western blot
analysis revealed that the expression levels of ZO-1 and Claudin-1
were significantly lower in the PH group than in the SO and PHP
groups. In addition, the plasma levels of D-lactic acid and endotoxin
and the amount of bacteria in the MLNs were significantly higher in
the PH group than in the other groups, indicating that the structure
of the intestines was tightly associated with the amount of damage
that was observed. Intestinal permeability was increased, and
intestinal bacteria and their products had penetrated the intestinal
epithelium and entered MLNs. This allowed them to invade other
tissues and organs, resulting in postoperative infection. This process
therefore represents one of the mechanisms by which bacterial
translocation can occur after major hepatectomy. The decreases in the
expression of ZO-1 and Claudin-1 that were observed in the cirrhotic
rats after major hepatectomy may have resulted from the following
underlying causes. a) An increase in portal pressure, which occurs
after major hepatectomy, causes the intestinal mucosa to exist in a
state of low perfusion. Intestinal epithelial cells therefore reside in a
state of severe hypoxia, leading to a locally harmful accumulation of
metabolites, which causes acidosis and stimulates the adherence,
aggregation and activation of white blood cells. These changes were
followed by the release of kinins, oxygen-free radicals, proteases, and
lysosomal enzymes. These factors cause the lipids in the cell and
mitochondrial membranes to undergo peroxidation and calcium
overload reactions, which eventually lead to cell death or epithelial
apoptosis. b) Increases in inflammatory factors may be involved
because the results of the present study show that the proportions of
cells that are strongly positive for TNF-α and IFN-γ expression were
significantly higher in the PH group than in the SO and PHP groups.
TNF-α is an important regulatory factor of intestinal mechanical
barrier function. A previous study showed that TNF-α induces
intestinal epithelial cells to undergo bulk shedding [16]. TNF-α upregulates
the expression of MLCK, as confirmed by our results [17-
19]. TNF-α induces NF-κB to translocate to the nucleus from the
cytosol, and factors downstream of TNF-α are then activated,
resulting in the promotion of apoptosis-related genes, such as Fas/
FasL genes, and the induction of epithelial cell apoptosis [20]. An
increase in TNF-α level can directly inhibit the expression of ZO-1
transcription [21]. IFN-γ promotes micropinocytosis in epithelial
cells, which results in the promotion of Claudin and Occlud in activity
and the subsequent failure of tight junction integrity [22]. c)
Endotoxin-damaged mitochondria and lysosomes were observed in
intestinal epithelial cells, and ischemia and hypoxia resulted in
shrinkage of intestinal vessels [23]. We found that endotoxin shad
translocated and entered the liver, where they stimulated Kupffer
cells to secrete a variety of inflammatory factors, resulting in a
continuous increase in the levels of inflammatory factors in the blood,
which caused damage to the intestinal mucosa [21]. d) MLCK
expression was up-regulated. Myosin undergoes conformational
changes when phosphorylated, resulting in the contraction of the
actin skeleton and leading to barrier dysfunction in the intestinal
epithelium. In addition, MLCK promotes the endocytosis of Occlud
in [18,19] in intestinal epithelial cells in response to inflammatory
mediators, such as TNF-α. e) Liver resection may have contributed to
the changes observed in their expression. After hepatectomy, the
number of cells in the reticuloendothelial system and the number of
Kupffer cells in the liver were significantly lower. The rate of hepatic
clearance of endotoxins and bacteria from the abdominal cavity
decreased in the liver, resulting in endotoxemia and bacteremia,
which in turn increased intestinal tight junction damage. The
intestinal microbial ecosystem plays an important role in the immune
system and the maturation of other physiological processes [24,25].
Under normal circumstances, various bacteria exist in a dynamic
equilibrium that constitutes the intestinal biological barrier. In the
present study, the mechanism underlying intestinal bacterial
translocation was associated with changes in the composition of fecal
bacteria and involved the following processes. a) Probiotic
antagonistic effects reduced pathogen colonization. Probiotics can
produce short-chain fatty acids that reduce intestinal PH and promote
the secretion of antimicrobial peptides [26]. The results of the present
study show that the amount of Bifidobacteria in the PH group was
only 69% of the amount observed in the SO group. This difference
benefited the reproduction of the pathogen Escherichia coli and
promoted damage to the intestinal epithelium [27]. In addition,
probiotic products, such as butyric acid, can up-regulate intestinal
epithelial cells via the antibacterial peptide LL-37, provide energy for
intestinal epithelial cells, and promote the proliferation and repair of
epithelial cells [28]. b) The levels of intestinal tight junction proteins
were decreased. Probiotics can up-regulate the expression of
Claudin-1 and Occludin-1, resulting in improved colonic permeability
[29], and inhibiting the expression of these proteins increases
intestinal permeability. Additionally, decreasing the amount of
Bifidobacteria increased cell membrane wrinkling by colonic
epithelial cells and down-regulated ZO-1 expression [30]. c) Intestinal
pathogenic bacteria were increased. There was a significant increase
in intestinal pathogenic bacteria in the PH group (i.e., Escherichia
coli, Clostridium, Klebsiella, and Bacteroides). When intestinal
bleeding occurs, Escherichia coli and Clostridium welchii can directly
damage or induce the phosphorylation and dephosphorylation of
closely connected structural proteins, resulting in the redistribution
of tight connection structures between the cell membrane and
cytoplasm and damaging their integrity [31]. d) Nutrient competition
is affected, in that probiotics reside in the human intestine, and
probiotics are better than pathogens at adapting to the intestinal
environment, and they show stronger survival skills and are better
able to compete for nutrients, which allows them to inhibit the growth
of pathogenic bacteria. In the present study, the number of
postoperative probiotics was lower, and the proportion of pathogenic
bacteria was higher, and these effects weakened the competitive
ability of the probiotic nutrients and caused the proportions of
maladjusted bacterial groups to continuously increase. e) A reduction
in intestinal probiotics damages the function of the intestinal immune
system. Unique types of bacteria that reside in the intestinal lumen
have been shown to adhere to the intestinal Peyer’s lymph nodes in
rats and to strongly stimulate regulatory T cells in the lamina propria,
resulting in an increase in the number of T cell subsets and there by
contributing to resistance in pathogen colonies and stimulating the
secretion of intestinal S-IgA [32,33]. f) Overgrowth of pathogenic
bacteria directly damages intestinal epithelial cells. The bacterial
proteases that are secreted by overgrown pathogenic bacteria were
found to damage microvilli by directly damaging microvilli
membrane proteins in intestinal epithelial cells [34].
The results of the present study show that after hepatectomy was
performed in cirrhotic rats, portal pressure increased, and the levels
of the inflammatory factors TNF-α and IFN-γ and the enzyme MLCK
were increased, resulting in damage to the intestinal mechanical
barrier. Additionally, in the rats that underwent surgery, the expression
of the ileal structural proteins ZO-1 and Claudin-1 was lower, the
composition of the intestinal bacteria was altered, the proportions
of probiotics were lower, the proportions of pathogens were higher,
and intestinal permeability had increased. The intestinal mechanical
and bacterial barriers were both damaged, and these effects represent
one of the mechanisms by which bacterial translocation can occur
after major hepatectomy. Propranolol can reduce portal pressure
and intestinal congestion, resulting in a reduction in damage to the
intestinal mucosal barrier, reducing endotoxemia, restoring balance
to and the maintenance of intestinal bacteria and reducing damage to
the intestinal mechanical barrier. Each of these effects may effectively
reduce bacterial translocation in the intestines.
Acknowledgement
This work was supported by the National Natural Science Foundation of China (grant number 81172783).
References
- Uchiyama K, Ueno M, Ozawa S, Kiriyama S, Kawai M, Hirono S, et al. Risk factors for postoperative infectious complications after hepatectomy. J Hepatobiliary Pancreat Sci. 2011;18(1):67-73.
- Capussotti L, Viganò L, Giuliante F, Ferrero A, Giovannini I, Nuzzo G. Liver dysfunction and sepsis determine operative mortality after liver resection. Br J Surg. 2009;96(1):88-94.
- Chen WZ, Hu KP, Xu RY, Pan WD. Beneficial effect of splenic artery ligation on bacterial translocation after major liver resection in rats. Dig Liver Dis. 2013;45(3):233–7.
- Ekberg H, Tranberg KG, Andersson R, Jeppsson B, Bengmark S. Major liver resection: perioperative course and management. Surgery. 1986;100(1):1-8.
- Kamada N, Calne RY. A surgical experience with five hundred thirty liver transplants in the rat. Surgery. 1983;93:64-69.
- Chiu CJ, McArdle AH, Brown R, Scott HJ, Gurd FN. Intestinal mucosal lesion in low-flow states. I. A morphological, hemodynamic, and metabolic reappraisal. Arch Surg. 1970;101(4):478-83.
- Wang X, Andersson R, Soltesz V, Bengmark S. Bacterial translocation after major hepatectomy in patients and rats. Arch Surg. 1992;127(9):1101-6.
- Wang XD, Pärsson H, Andersson R, Soltesz V, Johansson K, Bengmark S. Bacterial translocation, intestinal ultrastructure and cell membrane permeability early after major liver resection in the rat. Br J Surg. 1994;81(4):579-84.
- Nakao T, Kurita N, Komatsu M, Yoshikawa K, Iwata T, Utusnomiya T, et al. Irinotecan injures tight junction and causes bacterial translocation in rat. J Surg Res. 2012;173(2):341-7.
- Benten D, Wiest R. Gut microbiome and intestinal barrier failure--the “Achilles heel” in hepatology? J Hepatol. 2012;56(6):1221-3.
- McGhee JR, Fujihashi K. Inside the mucosal immune system. PLOS Biology. 2012.
- Watson AJ, Hughes KR. TNF-α-induced intestinal epithelial cell shedding: implications for intestinal barrier function. Ann N Y Acad Sci. 2012;1258:1-8.
- Nakagawa H, Tsunooka N, Yamamoto Y, Yoshida M, Nakata T, Kawachi K. Intestinal ischemia/reperfusion-induced bacterial translocation and lung injury in atherosclerotic rats with hypoadiponectinemia. Surgery. 2009;145(1):48-56.
- Van Itallie CM, Anderson JM. Claudins and epithelial paracellular transport. Annu Rev Physiol. 2006;68:403-29.
- Fasano A. Zonulin and its regulation of intestinal barrier function: the biological door to inflammation, autoimmunity, and cancer. Physiol Rev. 2011;91(1):151-75.
- Kiesslich R, Duckworth CA, Moussata D, Gloeckner A, Lim LG, Goetz M, et al. Local barrier dysfunction identified by confocal laser endomicroscopy predicts relapse in inflammatory bowel disease. Gut. 2012;61(8):1146-53.
- Ye D, Ma I, Ma TY. Molecular mechanism of tumor necrosis factor—a modulation of intestinal epithelial tight junction barrier. Am J Physiol Gastrointest Liver Physiol. 2006;290(3):G496-504.
- Shen L, Turner JR. Role of epithelial cells in initiation and propagation of intestinal inflammation. Eliminating the static: tight junction dynamics exposed. Am J Physiol Gastrointest Liver Physiol. 2006;290(4):G577-82.
- Utech M, Mennigen R, Bruewer M. Endocytosis and recycling of tight junction proteins in inflammation. J Biomed Biotechnol. 2010;2010:484987.
- Campbell KJ, Rocha S, Perkins ND. Active repression of antiapoptotic gene expression by RelA(p65) NF-kappa B. Mol Cell. 2004;13(6):853-65.
- Song HL, Lv S, Liu P. The roles of tumor necrosis factor-alpha in colon tight junction protein expression and intestinal mucosa structure in a mouse model of acute liver failure. BMC Gastroenterol. 2009;22:70.
- Bruewer M, Utech M, Ivanov AI, Hopkins AM, Parkos CA, Nusrat A. Interferon-gamma induces internalization of epithelial tight junction proteins via a macropinocytosis-like process. FASEB J. 2005;19(8):923-33.
- Catalioto RM, Maggi CA, Giuliani S. Intestinal epithelial barrier dysfunction in disease and possible therapeutical interventions. Curr Med Chem. 2011;18(3):398-426.
- Smith K, McCoy KD, Macpherson AJ. Use of axenic animals in studying the adaptation of mammals to their commensal intestinal microbiota. Semin Immunol. 2007;19(2):59-69.
- Hooper LV, Wong MH, Thelin A, Hansson L, Falk PG, Gordon JI. Molecular analysis of commensal host-microbial relationships in the intestine. Science. 2001;291(5505):881-4.
- Lupp C, Robertson ML, Wickham ME, Sekirov I, Champion OL, Gaynor EC, et al. Host-mediated inflammation disrupts the intestinal microbiota and promotes the overgrowth of Enterobacteriaceae. Cell Host Microbe. 2007;2(2):119-29.
- Keeney KM, Finlay BB. Enteric pathogen exploitation of the microbiota-generated nutrient environment of the gut. Curr Opin Microbiol. 2011;14(1):92-8.
- Maslowski KM, Vieira AT, Ng A, Kranich J, Sierro F, Yu D, et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature. 2009;461(7268):1282-6.
- Corridoni D, Pastorelli L, Mattioli B, Locovei S, Ishikawa D, Arseneau KO, et al. Probiotic bacteria regulate intestinal epithelial permeability in experimental ileitis by a TNF-dependent mechanism. PLOS ONE. 2012;7:0042067.
- Jones SE, Versalovic J. Probiotic Lactobacillus reuteri biofilms produce antimicrobial and anti-inflammatory factors. BMC Microbiol. 2009;9:35.
- Simonovic I, Rosenberg J, Koutsouris A, Hecht G. Enteropathogenic Escherichia coli dephosphorylates and dissociates occludin from intestinal epithelial tight junctions. Cell Microbiol. 2000;2(4):305-15.
- Gaboriau-Routhiau V, Rakotobe S, Lécuyer E, Mulder I, Lan A, Bridonneau C, et al. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity. 2009;31(4):677-89.
- Lara-Villoslada F, Sierra S, Boza J, Xaus J, Olivares M. Beneficial effects of consumption of a dairy product containing two probiotic strains, Lactobacillus coryniformis CECT5711 and Lactobacillus gasseri CECT5714 in healthy children. Nutr Hosp. 2007;22(4):496-502.
- Barber AE, Jones WG, Minei JP, Fahey TJ, Lowry SF, Shires GT. Bacterial over growth and intestinal at rophy in the etiology of gut barrier failure in the rat. Am J Surg. 1991;161(2):300-4.