Research Article
Sildenafil Transiently Delays Early Alveolar Bone Healing of Tooth Extraction Sockets
Elysse Orchard1, Wanda Green2, Renjith Parameswaran Nair3, Fleurette Abreo4 and Gulshan
Sunavala-Dossabhoy3*
1Department of Animal Resources, Louisiana State University Health Sciences Center, 1501 Kings Highway,
Shreveport, Louisiana 71130, USA
2Department of Cellular Biology and Anatomy, Louisiana State University Health Sciences Center, 1501 Kings
Highway, Shreveport, Louisiana 71130, USA
3Department of Biochemistry and Molecular Biology, Louisiana State University Health Sciences Center, 1501
Kings Highway, Shreveport, Louisiana 71130, USA
4Department of Pathology, Louisiana State University Health Sciences Center, 1501 Kings Highway, Shreveport,
Louisiana 71130, USA
*Corresponding author: Gulshan Sunavala-Dossabhoy, Department of Biochemistry and Molecular Biology, Louisiana State University Health Sciences Center, 1501 Kings Highway, Shreveport, Louisiana 71130, USA
Published: 03 May 2017
Cite this article as: Orchard E, Green W, Nair RP, Abreo
F, Sunavala-Dossabhoy G. Sildenafil
Transiently Delays Early Alveolar Bone
Healing of Tooth Extraction Sockets.
Clin Surg. 2017; 2: 1458.
Abstract
Bone is a unique tissue that has the ability to repair itself and return to full function. Bone regeneration
is a well-synchronized biological process that recapitulates embryonic bone development. The
establishment of a functional vascular supply has been shown to be essential for proper ossification
of newly deposited one, and impaired angiogenesis as in advanced age, diabetes, and anti-cancer
treatments affects bone repair. Endothelial guanosine 3', 5'-cyclic monophosphate (cGMP) is known
to support angiogenesis, and sildenafil, a phosphodiesterase 5 (PDE5) antagonist, prevents cGMP
hydrolysis and promotes the formation of new blood vessels. Since the development of functional
vascular networks is critical to bone repair, we investigated the effects of sildenafil on early alveolar
bone regeneration following exodontia. Our results demonstrate that per-oral administration of
sildenafil (10 mg/kg/day) in rats delays the dissolution and replacement of the sanguine clot with
granulation tissue. As a result, the number of replicating cells, a hallmark of regenerating tissue,
observed on day 4 is remarkably lower in sildenafil-treated animals than their control counterparts
(control: mean±SD, 47.35 ± 9.21; sildenafil: mean±SD, 11.47 ± 5.14). Similarly, cells expressing
transcription factor Cbfa-1/Runx2 and osteopontin, markers of differentiating osteoblasts, were
fewer in treated animals (control: mean±SD, 83.18 ± 4.60; sildenafil: mean±SD, 13.77 ± 4.63).
Treatment with hydrolysis-resistant cyclic GMP (cGMP) showed findings similar to sildenafiltreated
animals suggesting the negative impact of cGMP on early inflammatory phase of bone
healing. Nevertheless, histological differences were not significant between the 2 groups on day 8.
Based on these findings, we conclude that sildenafil transiently retards early events in alveolar bone
healing.
Keywords: PDE5, cGMP, Bone regeneration
Introduction
Bone regeneration is a biological process that involves a cascade of well-synchronized responses
essentially comprised of three overlapping stages namely, inflammatory, reparative, and remodeling.
Activation of endothelial cells and the sprouting of new capillaries from pre-existing vasculature is a
process referred to as angiogenesis. Angiogenesis is critical for bone healing [1,2], and angiogenesis
inhibitors have been shown to block the outgrowth of new blood vessels and delay the calcification of
newly deposited bone at fracture sites [3-5]. Embryonic development of facial bones and flat bones
of the skull occurs by intramembranous ossification, and bone regeneration within the alveolus after
tooth extraction recapitulates embryonic development. The fibrin clot formed soon after extraction
undergoes organization and is replaced with a stem cell-rich connective tissue. In response to
inflammatory signals, stem cells undergo differentiation to cells of angiogenic and osteogenic
lineages, and cellular events ultimately culminate in bone deposition and maturation. The formation
of new blood vessels is intimately connected to bone repair, and impaired angiogenesis as in old age,
diabetes, or radiotherapy delays bone healing.
The role of cGMP in modulating vascular tone and permeability during angiogenesis has been demonstrated before [6,7]. Phosphodiesterase 5 (PDE5) is a potent
enzyme that regulates intracellular cGMP levels by promoting the
conversion of cGMP to GMP. PDE5 antagonists such as sildenafil
suppress the hydrolysis of cGMP and increase the intracellular cGMP
pool. Sildenafil treatment has been shown to increase angiogenesis
and improve soft tissue survival after embolic stroke, ischemic limb
pathology, and random-pattern flap grafts in animals [8-11]. Nitric
Oxide (NO) interacts with soluble guanine cyclase to stimulate the
production of cGMP, and NO is known to improve tissue recovery
after ischemic pathology [12,13]. NO is a potent stimulator of
cell proliferation and angiogenesis during wound healing [14-
16], and wound repair is impaired and bone formation reduced in
endothelial nitric oxide synthase (eNOS)-deficient animals and in
animals administered nitric oxide synthase (NOS) inhibitors [17,18].
Moreover, osteoblasts of eNOS knockout animals demonstrated a
delay in bone maturation, whereas exogenous supplementation of
NO unregulated genes involved in osteoblast differentiation and
matrix mineralization [15]. Based on the involvement of NO/cGMP
pathway in bone regeneration, the aim of the study was to investigate
the effects of PDE5 inhibition on alveolar bone healing after tooth extraction.
Figure 1
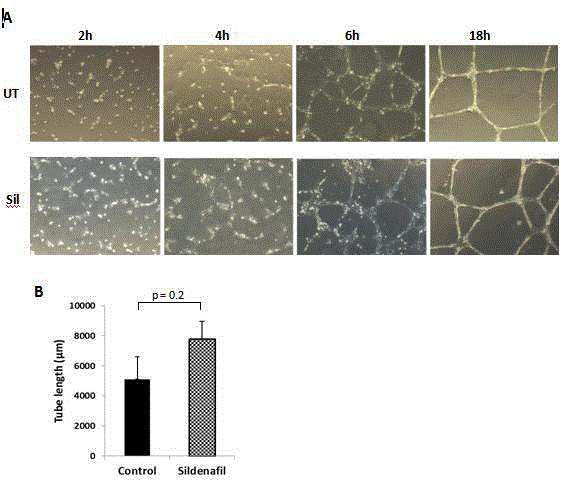
Figure 1
Sildenafil facilitates capillary tube formation in vitro. A) HUVEC cells plated on growth factor-reduced matrigel were treated, or not, with sildenafil (100
nM) and cells were imaged at 2, 4, 6, and 18 h. B) Quantification of total tube length at 4 h. Three random microscopic fields per well were imaged and total tube
length quantified by ImageJ Angiogenesis Analyzer. UT: untreated; Sil: sildenafil-treated
Figure 2
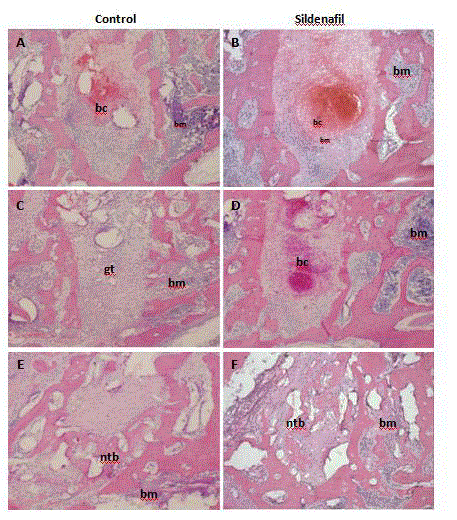
Figure 2
Blood clot resolution is delayed in response to sildenafil. Histological assessment of healing of extraction sockets at A,B) day 2 and C,D) day 4 in vehicle
treated controls (A,C) and sildenafil-treated animals (B,D). Images were captured at 40x magnification. bc: blood clot; bm: bone marrow; gt: granulation tissue.
Figure 3
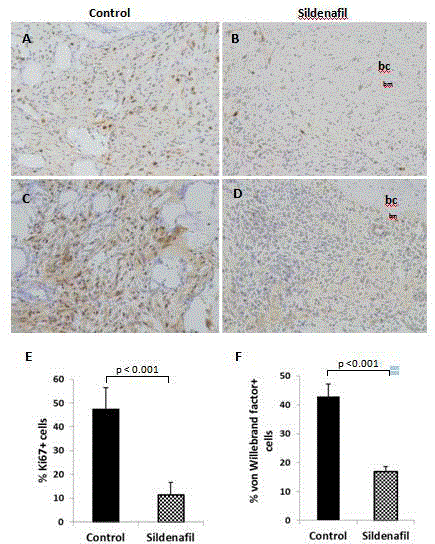
Figure 3
Sildenafil-associated decrease in cell proliferation and vascular density. Immunohistochemical detection of A,B) Ki-67 and C,D) von Willebrand factor
in decalcified, paraffin tissue sections of control and sildenafil-treated animals. E,F) Percent vascular density and proliferating cell population were measured as a
Materials and Methods
Reagents
Sildenafil was a kind gift from Pfizer (New York, NY), while
8-bromo-cGMP was purchased from Calbiochem. Primary
antibodies, anti-Ki67, anti-von Willebrand factor, anti-Osteopontin
and anti-Runx2 were purchased from Abcam. The HRP-conjugated
secondary antibodies and HRP-detection kit were obtained from
Biogenex.
Endothelial tube formation
Human umbilical vein endothelial cells, HUVEC, were
grown in Dulbecco’s Modified Eagles Medium (DMEM) that was
supplemented with 5% FBS, antibiotic-antimycotic (Invitrogen) and
minimal essential amino acids (Invitrogen). Capillary tube formation
was conducted in 6-well culture plates coated with low growth factor
matrigel (BD Biosciences). HUVEC cells were seeded at 1×104 cells/
cm2 in DMEM with 5% FBS with, or without, sildenafil (100 nM)
and cells were imaged at various times. Capillary tube formation
was quantified in 3 random areas of the well and the length of cords
between cells was measured using ImageJ Angiogenesis Analyzer.
Animals and Experimental procedure
Forty outbred male Wistar rats (Harlan/Envigo) weighing
approximately 250 gm were housed under 12 h dark and light cycle
with free access to food and water. Experiments were performed
under the aegis of the LSUHSC Animal Care and Use Committee,
and they conformed to the guidelines for the care and use of
laboratory animals. Animals were anesthetized with intramuscular
administration of ketamine chloride (42 mg/kg), xylazine (8 mg/
kg), and acepromazine (1.4 mg/kg), and the left mandibular first
molars were luxated. Bleeding was stanched by applying pressure
at the extraction site. Animals were kept warm during recovery and
were housed based on treatment. Sildenafil was dissolved in sterile
water, and 200 μl of sildenafil (10 mg/kg/day; divided into 2 doses
administered 10 h apart) or water was administered by gavage each
day starting soon after tooth extraction. Due to the 8-fold increase
in plasma clearance rate and a short half-life of sildenafil in rodents
(>1 hour in mice versus 4 hours in humans; [19, 20]), the dosage of
sildenafil was adjusted to achieve therapeutic dosing comparable
to humans. Animals were euthanized under anesthesia by carbon
dioxide inhalation on day 2, 4, or 8, and the left segment of the
mandible dissected and harvested. In a separate set of experiments,
8-bromo-cGMP (100 gm/kg), a cell-permeable non-hydrolysable
analogue of cGMP, was administered daily intravenously (n = 4).
Histology and Immunohistochemistry
The left segment of the mandible was dissected, and the bone
cleared of soft tissue before fixing in 4% paraformaldehyde/ phosphate
buffered saline (PBS), pH 7.4, for 48 h. Tissues were then decalcified
in 20% EDTA/ PBS, pH 7.4, for 7 days before paraffin processing and
embedding. Tissue sections were stained with hematoxylin-eosin for
histological evaluation, or used for immunohistochemistry. Antigen
retrieval of deparaffinized tissue sections was done by immersing
slides in DeCal Retrieval Solution (Biogenex), 20 m, followed by
incubation with anti-von Willebrand Factor (1:250), anti-Ki67
(1:100), anti-Runx2 (1:1000), or anti-Osteopontin (1:2500), 16 h at
4 oC. Tissue sections were washed and incubated in Biogenex HRPconjugated
universal secondary antibody for 30 m before reacting with diaminobenzidine (DAB) and counterstaining with hematoxylin.
Morphometric analysis of regenerating tissue
Slides stained with antibodies to Ki67 and von Willebrand
factor were used to measure cell proliferation and vascular indices.
Immunostaining in a total of 5 random fields was analyzed, and
proliferation index and vascular density were determined by percent
Ki67 + cells or von Willebrand factor + cells to the total number
of hematoxylin-stained nuclei. Immuno-reacted Runx2+ and
osteopontin+ cells were similarly measured to evaluate percent osteoangio-
progenitor cells and percent osteogenic cells. All data were
analyzed by Student's t-test to determine p-values.
Figure 4
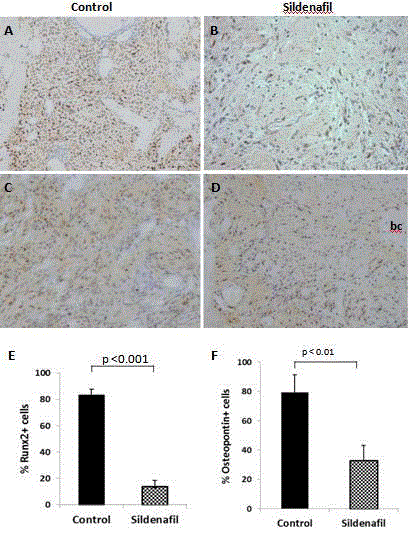
Figure 4
Decreased number of angio- osteo- progenitor cells after sildenafil treatment. Immuno-detection of A,B) Runx2 and C,D) Osteopontin in tissue samples
harvested on day 4. Control (A,C) and sildenafil-treated animals (B,D). Percent Runx2+ and Osteopontin+ immuno-stained cells in 5 random fields (200x
magnification).
Results
Sildenafil accelerates capillary tube formation in vitro
The effect of sildenafil on capillary tube formation was analyzed
after seeding HUVEC on growth factor-reduced matrigel and assessing
their organization at regular time intervals. Cells formed branching
networks within 6 h, which organized into tube-like structures by 18
h (Figure 1A). The addition of sildenafil hastened the development of
cell extensions and the formation of cell-cell connections. At 4 h, the
total length of continuous tube connections was greater in sildenafiltreated
cells than the non-treated counterparts (Figure 1B).
Sildenafil retards early bone healing
The establishment of blood supply is critical for proper bone
healing [5,21], and sildenafil was shown to accelerate fracture repair
in mice [22]. To evaluate whether the effect can be recapitulated
in alveolar bone healing, we assessed the early stages of healing of
molar extraction sockets over a period of 8 days. Hematoxylin and
eosin stained tissue sections of samples collected on days 2 and 4
following tooth extraction demonstrated a lengthier retention time of
the sanguine clot in sildenafil-treated animals compared to vehicletreated
(Figure 2A and B). A delay in clot resolution was also observed
after administration of non-hydrolysable analog of cGMP, 8-bromocGMP
(Supplementary Figure 1). A lag in infilling of granulation
tissue was the major difference observed between the groups (Figure
2C and D). However, in contrast to days 2 and 4, healing on day 8 was
comparable in drug-treated animals and controls (data not shown).
Delay in infiltration of stem/progenitor-rich connective
tissue in response to sildenafil
Granulation tissue composed of actively dividing and
differentiating stem/progenitor cells is a hallmark of tissue repair and
regeneration. Bone tissues harvested on day 4 demonstrated far fewer
cells that immuno-reacted with Ki67, a marker of proliferating cells,
in response to sildenafil treatment (control: mean ± SD, 47.35 ± 9.21;
sildenafil: mean ± SD, 11.47 ± 5.14; Figure 3A,B and E). Moreover,
an assessment of neovasculature by von Willebrand factor immunostaining
demonstrated a decrease in the number of immuno-reactive
cells following drug treatment (control: mean ± SD, 42.9 ± 4.3;
sildenafil: mean ± SD, 17.0 ± 1.8; Figure 3C,D and F).
Essential to early osteoblast differentiation is the expression of
osteoblast-specific transcription factor, Runt-related Transcription
factor 2 (Runx2). Runx2 is highly expressed in immature osteoblasts,
but is downregulated during maturation [23]. However, Runx2
is not an exclusive marker of osteogenic cells. It is expressed in endothelial progenitor cells too and has been found to be important
for endothelial cell programming [24,25]. To assess the contribution
of osteo- angio-progenitor cell population to bone healing, tissue
sections were incubated with anti-Runx2. Quantitative morphometric
measurement from 5 random fields demonstrated a near 6-fold
decrease in cell-staining after drug treatment. (control: mean ± SD,
83.18 ± 4.60; sildenafil: mean ± SD, 13.77 ± 4.63; (Figure 4A,B and E).
Sildenafil retards bone matrix deposition
Runx2 induces the expression of important bone matrix
proteins namely, osteopontin, bone sialoprotein, osteocalcin and
Col1α1 in immature osteoblasts and mesenchymal progenitors
during bone development [23]. To analyze whether the decrease in
Runx2-immunoreactive cells translates to a decrease in osteopontin
expression after sildenafil treatment, osteopontin immunolocalization
was conducted on tissues collected at day 4. As expected,
the results demonstrated that exposure to the drug results in fewer
osteopontin-expressing cells (control: mean ± SD, 79.58 ± 11.81;
sildenafil: mean ± SD, 33.12 ± 10.17; Figure 4C,D and F).
Supplementary Figure 1
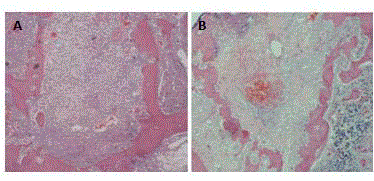
Supplementary Figure 1
Increase in cGMP signaling delays blood clot
remission. Hematoxylin–eosin staining of bone isolated from A) control and
B) 8-bromo-cGMP treated animals 4 days after tooth extraction.
Discussion
Angiogenesis and the establishment of a functioning vascular bed
have been shown to be vital to successful bone repair. Angiogenesis
agonists such as vascular endothelial growth factor (VEGF) stimulate
bone repair [26], whereas angiogenesis inhibitors suppress bone
healing at fracture and implant sites [3-5]. PDE5 is the most active
of all cGMP-hydrolyzing PDEs, and it is widely expressed in tissues
including bone [27]. In light of previous studies that demonstrated
PDE5 antagonism in promoting neovascularization through
augmentation of endothelial cGMP and increasing in bone marrowderived
endothelial progenitor cells [27], we anticipated a positive
impact of PDE5 inhibitor, sildenafil, on early bone healing. Similar to
previous studies, we demonstrated that sildenafil increases HUVEC
migration and formation of capillary-like structures on matrigel in
vitro [13,19]. Angiogenesis and early alveolar bone healing when
assessed in vivo showed that the sequential phases of healing of
extraction sockets described before were reproducible in our study [28].
However, the addition of sildenafil retarded blood clot remission and
the migration of stem cells into the defect. Since sildenafil principally
acts through the cGMP signaling pathway a delay in the resolution
of the fibrin clot in cell-permeable 8-bromo-cGMP treated animals
implicated cGMP in attenuating clot dissolution. Differentiation
of monocytes to macrophages or dendritic cells is induced by
inflammatory stimuli, and evidence indicates that cyclic nucleotides
such as cGMP can suppress the process [29]. Our results suggest that
sildenafil-initiated increase in cGMP negatively influences early bone
healing. Alveolar bone formation after tooth extraction occurs by the
direct differentiation of mesenchymal stem cells to cells of osteogenic
lineage. The important role of Runx2 in osteogenesis was confirmed
when Runx2 knockout mice displayed poor skeletal development
[30,31]. Runx2 is also expressed in endothelial precursors and it
regulates their migration, proliferation, and differentiation [24,32].
Runx2 is, therefore, expressed in cells committed to osteogenic
and angiogenic fates, and our s suggest that recruitment of stem/
progenitor cells to the site of repair is impaired by sildenafil. Earlier
studies investigating sildenafil were focused on angiogenic outcomes,
and only recently have studies noted its anti-inflammatory effects
[33-36]. Inflammation is a critical initial event in a tissue’s response to
injury. It signals damage and activates tissue repair mechanisms. The
inflammatory response initiated cascade of events plays a significant
role in bone repair, and its suppression delays healing [37-40]. A
recent study examining the effects of sildenafil on long bone fracture
repair found that it negatively affects the inflammatory phase of bone
healing [41]. Our findings corroborate the finding that the drug
attenuates early events in bone healing. It is suggested that the antiinflammatory
effects of sildenafil suppress blood clot resolution and
delays, the migration of mesenchymal stem cells to the site of injury.
Previous studies that analyzed long bone fracture repair at >2 weeks
have demonstrated a beneficial effect of sildenafil [22,41], and with
no appreciable differences in healing observed at day 8, we suggest
that the drug accelerates reparative phase of alveolar bone healing
presumably through improved angiogenesis.
The preservation and maintenance of adequate bone volume after
tooth extraction improves the success of prosthetic rehabilitation.
Our findings of an early, but transitory, delay in bone healing by
sildenafil suggest that a prudent measure would be to avoid the drug
in the days immediately following tooth extraction or placement of
dental implants.
Acknowledgements
The authors thank Senthilnathan Palaniyandi for his contribution during the early stages of the capillary tube formation study. The research was supported by funds from the American Cancer Society, National Institutes of Health: NIH/NCI R21CA173162, and the Feist- Weiller Cancer Center.
References
- Hankenson KD, Dishowitz M, Gray C, Schenker M. Angiogenesis in bone regeneration. Injury. 2011;42(6):556-61.
- Stegen S, van Gastel N, Carmeliet G. Bringing new life to damaged bone: the importance of angiogenesis in bone repair and regeneration. Bone. 2015;70:19-27.
- Hausman MR, Schaffler MB, Majeska RJ. Prevention of fracture healing in rats by an inhibitor of angiogenesis. Bone. 2001;29(6):560-4.
- Holstein JH, Klein M, Garcia P, Histing T, Culemann U, Pizanis A, et al. Rapamycin affects early fracture healing in mice. Br J Pharmacol. 2008;154(5):1055-62.
- Mair B, Fuerst G, Kubitzky P, Tangl S, Bergmeister H, Losert U, et al. The anti-angiogenic substance TNP-470 impairs peri-implant bone formation: a pilot study in the rabbit metaphysis model. Clin Oral Implants Res. 2007;18(3):370-5.
- Kemp-Harper B, Schmidt HH. cGMP in the vasculature. Handb Exp Pharmacol. 2009;191:447-67.
- Weyerbrock A, Walbridge S, Saavedra JE, Keefer LK, Oldfield EH. Differential effects of nitric oxide on blood-brain barrier integrity and cerebral blood flow in intracerebral C6 gliomas. Neuro Oncol. 2011;13(2):203-11.
- Lian Li, Quan Jiang, Li Zhang, Guangliang Ding, Zheng Gang Zhang, Qingjiang Li, et al. Angiogenesis and improved cerebral blood flow in the ischemic boundary area detected by MRI after administration of sildenafil to rats with embolic stroke. Brain Res. 2007;1132(1):185-92.
- Senthilkumar A, Smith RD, Khitha J, Arora N, Veerareddy S, Langston W, et al. Sildenafil promotes ischemia-induced angiogenesis through a PKG-dependent pathway. Arterioscler Thromb Vasc Biol. 2007; 27(9): 1947-54.
- Ulusoy MG, Uysal A, Koçer U, Karaaslan O, Cuzdan SS, Ayyildiz A, et al. Improved flap viability with site-specific delivery of sildenafil citrate using fibrin glue. Ann Plast Surg. 2005;55(3):292-6.
- Zhang L, Zhang RL, Wang Y, Zhang C, Zhang ZG, Meng H, et al. Functional recovery in aged and young rats after embolic stroke: treatment with a phosphodiesterase type 5 inhibitor. Stroke. 2005;36(4):847-52.
- Kumar D, Branch BG, Pattillo CB, Hood J, Thoma S, Simpson S, et al. Chronic sodium nitrite therapy augments ischemia-induced angiogenesis and arteriogenesis. Proc Natl Acad Sci USA. 2008;105(21):7540-5.
- Zhang R, Wang L, Zhang L, Chen J, Zhu Z, Zhang Z, Chopp M. Nitric oxide enhances angiogenesis via the synthesis of vascular endothelial growth factor and cGMP after stroke in the rat. Circ Res. 2003;92(3):308-13.
- Luo JD, Chen AF. Nitric oxide: a newly discovered function on wound healing. Acta Pharmacol Sin. 2005;26(3):259-64.
- Afzal F, Polak J, Buttery L. Endothelial nitric oxide synthase in the control of osteoblastic mineralizing activity and bone integrity. J Pathol. 2004;202(4):503-10.
- Diwan AD, Wang MX, Jang D, Zhu W, Murrell GA. Nitric oxide modulates fracture healing. J Bone Miner Res. 2000;15(2):342-51.
- Aguirre J, Buttery L, O'Shaughnessy M, Afzal F, Fernandez de Marticorena I, Hukkanen M, Endothelial nitric oxide synthase gene-deficient mice demonstrate marked retardation in postnatal bone formation, reduced bone volume, and defects in osteoblast maturation and activity. Am J Pathol. 2001;158(1): 247-57.
- Lee PC, Salyapongse AN, Bragdon GA, Shears LL 2nd, Watkins SC, Edington HD, et al. Impaired wound healing and angiogenesis in eNOS-deficient mice. Am J Physiol. 1999;277(4 Pt 2):H1600-8.
- Walker DK, Ackland MJ, James GC, Muirhead GJ, Rance DJ, Wastall P, et al. Pharmacokinetics and metabolism of sildenafil in mouse, rat, rabbit, dog and man. Xenobiotica. 1999;29(3):297-310.
- Halaris AE, Belendiuk KT, Freedman DX. Antidepressant drugs affect dopamine uptake. Biochem Pharmacol. 1975;24(20):1896-7.
- Warrington JS, Von Moltke LL, Shader RI, Greenblatt DJ. In vitro biotransformation of sildenafil (Viagra) in the male rat: the role of CYP2C11. Drug Metab Dispos. 2002;30(6):655-7.
- Cardaropoli G, Araújo M, Lindhe J. Dynamics of bone tissue formation in tooth extraction sites. An experimental study in dogs. J Clin Periodontol. 2003;30(9):809-18.
- Histing T, Marciniak K, Scheuer C, Garcia P, Holstein JH, Klein M, et al. Sildenafil accelerates fracture healing in mice. J Orthop Res. 2011;29(6):867-73.
- Maruyama Z, Yoshida CA, Furuichi T, Amizuka N, Ito M, Fukuyama R, et al. Runx2 determines bone maturity and turnover rate in postnatal bone development and is involved in bone loss in estrogen deficiency. Dev Dyn. 2007;236(7):1876-90.
- Sun L, Vitolo M, Passaniti A. Runt-related gene 2 in endothelial cells: inducible expression and specific regulation of cell migration and invasion. Cancer Res. 2001;61(13):4994-5001.
- Zelzer E, Glotzer DJ, Hartmann C, Thomas D, Fukai N, Soker S, et al. Tissue specific regulation of VEGF expression during bone development requires Cbfa1/Runx2. Mech Dev. 2001;106(1-2):97-106.
- Keramaris NC, Calori GM, Nikolaou VS, Schemitsch EH, Giannoudis PV. Fracture vascularity and bone healing: a systematic review of the role of VEGF. Injury. 2008;39(2):S45-57.
- Foresta C, Caretta N, Lana A, De Toni L, Biagioli A, Vinanzi C, Ferlin A. Relationship between vascular damage degrees and endothelial progenitor cells in patients with erectile dysfunction: effect of vardenafil administration and PDE5 expression in the bone marrow. Eur Urol. 2007;51(5):1411-7.
- Kanyama M, Kuboki T, Akiyama K, Nawachi K, Miyauchi FM, Yatani H, et al. Connective tissue growth factor expressed in rat alveolar bone regeneration sites after tooth extraction. Arch Oral Biol. 2003; 48(10):723-30.
- Giordano D, Magaletti DM, Clark EA, Beavo JA. Cyclic nucleotides promote monocyte differentiation toward a DC-SIGN+ (CD209) intermediate cell and impair differentiation into dendritic cells. J Immunol. 2003;171(12): 6421-30.
- Takarada T, Hinoi E, Nakazato R, Ochi H, Xu C, Tsuchikane A, et al. An analysis of skeletal development in osteoblast-specific and chondrocyte-specific runt-related transcription factor-2 (Runx2) knockout mice. J Bone Miner Res. 2013;28(10): 2064-9.
- Takarada T, Nakazato R, Tsuchikane A, Fujikawa K, Iezaki T, Yoneda Y, et al. Genetic analysis of Runx2 function during intramembranous ossification. Development. 2016;143(2): 211-8.
- Qiao M, Shapiro P, Fosbrink M, Rus H, Kumar R, Passaniti A. Cell cycle-dependent phosphorylation of the RUNX2 transcription factor by cdc2 regulates endothelial cell proliferation. J Biol Chem. 2006;281(11):7118-28.
- de Visser YP, Walther FJ, Laghmani el H, Boersma H, van der Laarse A, Wagenaar GT. Sildenafil attenuates pulmonary inflammation and fibrin deposition, mortality and right ventricular hypertrophy in neonatal hyperoxic lung injury. Respir Res. 2009;10:30.
- Di Luigi L, Corinaldesi C, Colletti M, Scolletta S, Antinozzi C, Vannelli GB, et al. Phosphodiesterase Type 5 Inhibitor Sildenafil Decreases the Proinflammatory Chemokine CXCL10 in Human Cardiomyocytes and in Subjects with Diabetic Cardiomyopathy. Inflammation. 2016;39(3):1238-52.
- Nunes AK, Rapôso C, Luna RL, Cruz-Höfling MA, Peixoto CA. Sildenafil (Viagra(R)) down regulates cytokines and prevents demyelination in a cuprizone-induced MS mouse model. Cytokine. 2012;60(2):540-51.
- Vlachopoulos C, Ioakeimidis N, Rokkas K, Angelis A, Terentes-Printzios D, Stefanadis C, et al. Acute effect of sildenafil on inflammatory markers/mediators in patients with vasculogenic erectile dysfunction. Int J Cardiol. 2015;182:98-101.
- Simon AM, Manigrasso MB, O'Connor JP. Cyclo-oxygenase 2 function is essential for bone fracture healing. J Bone Miner Res. 2002;17(6):963-76.
- Barthel W, Markwardt F. Aggregation of blood platelets by adrenaline and its uptake. Biochem Pharmacol. 1975;24(20):1903-4.
- Yugoshi LI, Sala MA, Brentegani LG, Lamano Carvalho TL. Histometric study of socket healing after tooth extraction in rats treated with diclofenac. Braz Dent J. 2002;13(2):92-6.
- Schmidt-Bleek K, Schell H, Schulz N, Hoff P, Perka C, Buttgereit F, et al. Inflammatory phase of bone healing initiates the regenerative healing cascade. Cell Tissue Res. 2012;347(3):567-73.
- Takayanagi H. Osteoimmunology: shared mechanisms and crosstalk between the immune and bone systems. Nat Rev Immunol. 2007;7(4):292-304.
- Kilinc CY, Selahattin O, Acar E, Baris K. Effects of sildenafil on the inflammatory and repair phase of bone healing speed in a rat model. Acta Medica Mediterranea. 2015;31:1203-8.