Review Article
Photodynamic Therapy as a Treatment to Nasopharyngeal Carcinoma
WU Wing Kei Ricky1*, CHU SM Ellie1 and Yow MN Christine2
1Department of Medical and Health Sciences, Tung Wah College, China
2Department of Laboratory Sciences, Hong Kong Polytechnic University, China
*Corresponding author: WU Wing Kei Ricky, Department of Medical and Health Sciences, Tung Wah College, Hong Kong, China
Published: 13 Feb, 2017
Cite this article as: WU Wing Kei Ricky, CHU SM Ellie, Yow
MN Christine. Photodynamic Therapy
as a Treatment to Nasopharyngeal
Carcinoma. Clin Surg. 2017; 2: 1308.
Abstract
NPC is highly prevalent in Hong Kong with more than 800 new cases reported annually. The
epidemiologic evidence implies that Epstein-Barr virus (EBV) infection, environmental factors and
genetic factors play roles in the tumorigenesis of NPC. Conventional treatment of NPC is chemoradiotherapy,
yet the treatment outcomes in patients with advanced stage of NPC are unsatisfactory.
Therefore development of new treatment strategies is crucial for patients with Nasopharyngeal
Carcinoma (NPC), particularly for patients with drug resistance properties or distance recurrence.
Photodynamic therapy (PDT) is an FDA approved cancer regimen which employs a combination
of light-activated photosensitiser visible light and molecular oxygen to selectively destroy the
biological targets. 3rd generation of photosensitizers with light activation by advance light sources
such as coherent light source (laser) and non-coherent light source (LED) could be one of novel
strategies in for NPC patients. In-depth investigation for selected PSs mediated PDT on NPC model
is still underway.
Keywords: Photodynamic therapy; Nasopharyngeal carcinoma; Epstein barr virus; Microrna; Drug resistance
Introduction
Overview of nasopharyngeal carcinoma (NPC)
Nasopharyngeal carcinoma (NPC) is endemic in Asia and is one of the top ten cancers highly
prevalent in Hong Kong. The overall incidence is 6.5/100,000 person-years in southeastern Asia. Yet
the incidence rate increase sharply in Hong Kong to12.2/100,000 person-years [1-4]. Nasopharyngeal
Carcinoma (NPC) encompasses any squamous cell carcinoma arising in the epithelial lining of the
nasopharynx and is characterized by poor or undifferentiated carcinoma [5,6].
Classification of NPC
The histopathological classification of NPC was distinguished into three types by The World
Health Organization according to the degree of differentiation. Type I is keratinizing squamouscell
carcinoma similar to carcinomas that arise from other sites of the head and neck. Type II is
non-keratinizing epidermoid carcinomas. Type III represents the undifferentiated carcinomas [2,7].
Among these, NPC Type I is uncommon in endemic areas while type II and type III NPC are more
common and are closely related to EBV infection [8].
NPC can be also staged clinically according to the 7th edition of the International Union against
Cancer (UICC) and the American Joint Committee on Cancer (AJCC) staging-system manual.
AJCC classifies NPC into IV stages according to the TNM system. TNM system contains 3 key
pieces of information, includes T) describes whether the primary tumor has invaded into nearby
tissues or organs, N) describes whether the primary tumor has spread to nearby lymph nodes, and
M) describes whether the cancer has metasized. Different score are given to the TNM system in
order to provide more details about each of these descriptions. The number 2 to 4 indicate the
degree of spread, while X will be given to those cases which cannot be assessed. Stage I NPC is T1-
N0-M0, indicates tumor is in the nasopharynx and may spread to nearby soft tissues. Stage II NPC
is T2-N0-M0 or T1/T2-N1-M0, indicated the tumor has spread into nearby tissue. Stage III NPC is
T3-N0 to N2-M0 or T1/T2-N2-M0, indicates the tumor has spread to lymph nodes. Stage IV NPC
is T4-N0 to N2-M0 or Any T-N3-M0 or Any T-Any N-M1, indicates the tumor may have spread
to distant sites [9].
NPC Tumorigenesis
It is widely accepted that Epstein-Barr virus (EBV) infection plays
a major role in the tumorigenesis of NPC. EBV is a herpes virus that
infects over 90% of adult population and is consistently detected
in NPC patients [10,11]. Studies reported that EBV is one of the
most potent transforming agents for human cells and is associated
with a number of malignancies, including Burkett’s lymphoma
and nasopharyngeal carcinoma [12-14]. It is rare to observe viral
replication in EBV-infected cells as EBV establishes a latent infection
with a restricted set of latent gene being expressed, including two
EBV-encoded nuclear RNAs (EBER1, EBER2), six EBV-encoded
nuclear antigens (EBNA1, EBNA2, EBNA3A, EBNA3B, EBNA3C,
EBNA-LP), and three latent membrane proteins (LMP1, LMP2A,
LMP2B). These latent gene expression is now identified at three
patterns, which is known as latency I, II and III. Only the EBERs and
ENBA1 are expressed in latency I; the EBERs, EBNA1, LMP1 and
LMP2 are expressed in latency II; and all latent genes are expressed in
latency III [12,15-19].
In NPC, EBV replicates and hides in cells followed by type II
latency infection cycle, with expression of a limited number of viral
proteins, including the latent membrane proteins (LMP1, LMP2A
and LMP2B) and EBV-determined nuclear antigens (EBNA1 and
EBNA2).Among these, LMP1 is the principal oncogene involves in
the process of EBV-associated oncogenesis of NPC [17,20-26]. LMP1
is a 66kDa integral membrane protein consists of a 6 transmembrane
domains and a carboxyl-terminus containing 3 signaling domains
called C-terminal activating regions 1, 2 and 3 (CTAR 1, CTAR 2
and CTAR 3). The three CTAR domains provide docking sites for
signaling adaptor proteins. Among these, CTAR 1 and CTAR 2 are
two of the distinct functional domains responsible for the possess of
most of the LMP1 signaling activity via directly activate a number
of signaling pathways including nuclear factor kappa B (NF-kB),
mitogen-activated protein kinases (MAPK) and Janus Kinase/
Signal Transducer and Activator of Transcription (JAK/STAT)
pathway [8,23,27,28]. The LMP1 induced signal pathways is one of
the elemental factors controlling the biological behaviors of NPC.
Signaling pathways involved are mainly control cell functions such as
the inhibition of apoptosis; induction of cell immortality; promotion
of cell proliferation and influence the cell invasion and metastasis
[7,16,29,30].
EBV Encoded Micro RNAs
In additional to the well-established viral protein expression,
EBV has been found to express various RNAs. These have been
referred to as complementary strand transcripts (CSTs), BamHI A
rightward transcripts (BARTs) or the BARF0 RNAs [31,32]. The
possible role of these RNAs may affect viral transformation as high
levels of CST and BARTs expression have been detected in NPC
tumors [20,32,33]. The recent discovery of BART micro RNAs
(miRNA) has shed new light on the function of these transcripts.
miRNAs are a class of 19-24 nucleotide non-coding RNAs which
modulate gene expression. miRNAs are produced by endogenous
enzymatic (Dicer) digestion of RNA transcripts containing hairpin.
Protein translation is inhibited by forming complementary duplexes
of miRNA with their target mRNAs and cause degradation of these
mRNAs [29,34]. The number of EBV miRNAs made up 23.2% of the
total miRNAs in the biopsy samples in NPC patients, whereas only
0.1% EBV miRNAs is found in adjacent normal nasopharynx tissue
[35]. These potent gene regulators are through to control a wide range
of biological functions, including differentiation, cell growth and
cancer development [34,36,37]. Several studies reveal the function
of EBV encoded miRNAs to modulate both viral and cellular gene
expression in NPC cells, alternating the anti-apoptotic function,
immune evasion, and viral protein expression patterns. MiR-BART5-
5p was found to be down regulated the expression of a pro-apoptotic
protein, PUMA in C666-1 cells. It can keep the cell survive and
thus the period of latency increase [38]. miR-BART16 could down
regulate TOM22, a mitochondrial receptor for the pro-apoptotic
protein Bax. miR-BART2-5p could help the host cell to escape from
NK cell recognition by repressing the expression of cellular stressinduced
immune molecule (MICB). miR-BART 3 targets IPO7, a
nuclear importer receptor which has been showed to be responsible
for the nuclear translocation of transcription factor and is involved
in cytokine and early gene expression in activated T cells. miRBART6-
5p could suppress viral onco-protein EBNA2, preventing the
transition from latency I and II to III [39]. miR-BART2-5p can inhibit
EBV lytic replication by targeting the viral DNA polymerase BALF5.
miR-BART22 can down regulate the expression of LMP2A. It can
inhibit telomerase reverse transcription and induce anti-proliferation
by NF-kappa B suppression.
EBV miRNAs demonstrate differential expression in different
latent stages. EBV BARTs produce two clusters of miRNA and that
the BART Cluster 1 miRNAs target the LMP1 protein expression
and BART Cluster 2 miRNAs target EBV DNA polymerase BALF5
for degradation, effectively inhibiting lytic replication and help
EBV establish a type II latency in infected cells [29,36,38,40]. EBV
constitutively express up to 44 mature miRNAs and their main target
is its oncogene LMP1 [29]. Recent report indicate three BARTs cluster
1 miRNAs (ebv-miR-BART1-5p, 16 and 17-5p) targeting the LMP1
gene and down regulating the LMP1 protein expression [27,34,41].
One possible reason for the high level expression of miRNAs in EBV
is to avoid over expression of LMP1 protein. The over expression
of LMP1 protein could results in the inhibition of cell proliferation
and increase in apoptosis ref. Therefore, introducing miRNAs could
inhibit excessive MP1 expression in NPC cells and results in resistant
to the apoptosis [27,34,41-45].
Drug Resistance Mechanisms in NPC
Multidrug resistance is the major obstacle to chemotherapy in tumor patients. Development of MDR may be intrinsically prior to treatment or acquired during treatment [46]. Increase drug efflux from the cells via the adenosine triphosphate (ATP) binding cassette transporters (ABC), inactivation of drugs via detoxifying enzymes, and defective apoptotic pathways are three most common cause of MDR development [47,48]. Among these, the increase drug efflux via ABC membrane transporters is one of the leading mechanisms of MDR development in tumor cells [49,50]. ABC membrane transporters includes P-glycoprotein (P-gp/ABCB1), multidrug resistance associated protein 1 (MRP-1/ABCC1) and breast cancer resistance protein (BCRP/ABCG2). Among these, P-gp is the best studied mechanisms of MDR phenotype [51,52].
Multidrug Resistance Mechanisms Developed in Nasopharyngeal Carcinoma
There are only few articles reported the correlation of multidrug
resistance protein expression on EBV infected NPC cells. A few studies
demonstrated that P-gp and MRP1 were found to be expressed in NPC cells but in different levels. Study has shown that a small portion
of NPC express MDR1. Study demonstrated a 12.6% NPC patients
and 32.6% recurrent NPC patients expressed MDR1 [53]. Another
group of researcher reported that only 3.3% of tested NPC patients
expressed MDR1 [54]. Study also reported a significantly higher
expression level of MDR1 in keratinizing squamous cell carcinoma
type NPC than those with non-keratinizing and undifferentiated
types NPC. MDR1 is expressed at the apical surface of normal
nasopharyngeal epithelial cells, which protects normal tissue against
exogenous toxins and hydrophobic xenobiotics.
Different from MDR1 expression, the expression of MRP1 varied
among the NPC samples. Study reported that 40% of the tested NPC
patient samples expressed MRP. The high expression rate of MPR is
proofed to be correlated with the clinical stage. The MRP1 expression
could be one of the prognostic markers at the time of diagnosis
before treatment. However, study fails to illustrate the correlation
between MRP and chemo-sensitivity testing with selected anticancer
drugs, such as CDDP, 5-FU, PEP, MMC and ADM. Furthermore,
concerning the overall 5-year survival rate, there was no difference
between NPC exhibiting MRP and NPC not exhibit MRP [55].
Similar result was obtained by another group of researcher [54].
ABCB1/P-glycoprotein (P-gp)
Phenomenon of multidrug resistance was initially noticed in 1948 in leukemia patients by Farber [56]. In 1976, Ling’s group reported that the MDR phenotype is related to decreased intracellular drug accumulation mediated by a 170kDa plasma membrane glycoprotein, which known as the P-glycoprotein [57]. The association of high MDR1 expression levels with intrinsically resistant cancers and the increased expression of the MDR1 gene with cancers acquired drug resistance during treatment confirm the important role of MDR1 in human cancers. Recent studies indicated that P-gp is the most typical ATP-dependent drug efflux pump contributes to multidrug resistance in cancer cells and is the best-characterized mechanism of MDR. P-gp is a 170kDa trans membrane protein consists of two hydrophobic trans membrane domains (TMDs), three membrane spanning domain (MSD) and two nucleotide binding domain (NBDs). It is encoded by MDR1 gene and is located on chromosome 7. [58,59]. The TMDs region that form a pore-like structure contains the drug binding sites for different drugs while the NBDs responsible for the ATP binding and hydrolysis that drives drug transport [60,61].
Mechanism of Action of P-gp
Drug binging to the pore-like structure (high-affinity drug binding site) initiates the transport cycle via binding and hydrolysis of ATP coupled with P-gp. ATP is bound to nucleotide binding domains leads to conformational change of P-gp. Changes in P-gp conformation results as reduced affinity for drug binding and reorientation of the site so that it is exposed to the extracellular medium [60,62,63]. Studies have also shown that the binding of ATP, rather than hydrolysis of ATP, provide the energy for drug translocation [59,64-66]. The hydrolysis of ATP and release of ADP from P-gp does affect its conformation, returns the drug binding site to high drug affinity status.
Conventional Treatment of NPC
Treatment selected for NPC patients were based on the AJCC classification system. The conventional treatment for NPC is chemoradiotherapy because of high radio- and chemo-sensitivity with a 5-years overall survival of 70-80% for stage I and II NPC. Unfortunately majority of NPC patients were diagnosed with locally advanced stages as it is difficult to detect early because of its complex anatomical location [7,67]. The treatment outcomes in patients with stage II NPC become less favour that with stage I NPC because of the distance recurrence. Local recurrence, distant recurrence and development of multi-drug resistance properties are the most common cause of treatment failure [8,9, 54]. Complications always resulted after cheom-radiotherapy, such as hearing impairment, endocrinological dysfunctions, temporal lobe necrosis, cranial neuropathy, haemorrhage, and bone necrosis [68,69]. Thus development of new treatment strategies is crucial for patients with NPC, particularly for those patients with multi drug resistance properties.
Overview of Photodynamic Therapy (PDT)
Photodynamic therapy (PDT) is an evolving cancer treatment regimen with approved for use in USA, EU, Canada, Russia and Japan [70-72]. PDT uses a combination of photosensitizing agents (PS), visible light and molecular oxygen to selectively destroy the biological target. None of these is individually toxic, but together they initiate photo-destruction to biological target. In general, tumour localizing photosensitizers will absorb photon to produce photo-toxin such as singlet oxygen (1O2) and reactive oxygen species (ROS). Depends on the cellular organelles where PSs localized, ROS generated could oxidize many biological molecules, such as protein, lipids and nucleic acids and lead to in vivo and in vitro tumour cell disruption through apoptosis, necrosis and autophagy [73,74]. The antitumor effects of PDT derive from 3 mechanisms; including direct cytotoxicity effects on tumour cells, destruction of tumour associated vasculature, and induction of inflammatory reaction against tumour cells [70,73-84].
Photosensitizers
The prerequisites of an ideal photosensitizer including: chemical purity, low dark toxicity, high quantum yield of singlet oxygen, selective accumulation in tumour cells, short time interval between drug administration and maximal accumulation within target cells, rapid clearance from the body, and being activated by longer wavelength with better tissue penetration [85].
Hematoporphyrin Derivative (HpD) – Photofrin
The first FDA approved photosensitizer was Hematoporphyrin derivative (HpD). It was developed in the 1970s and early 1980s and is now known as the 1st generation photosensitizers for the treatment of tumor, such as early and late endobronchial lesions, Barrett’s esophagus and esophageal obstructing lesions with high response rate and promising result obtained [86-89]. It is a mixture of monomers, dimmers and oligomers of hematoporphyrin synthesized by chemical manipulation and are the photosensitizer that brought PDT to a worldwide audience. The first report of the preparation of HpD was published by Dougherty in 1983[90]. Afterward, a number of HpD formulas were available in commercial and Photofrin from Axcan Pharma was the one commonly used for tumor treatment. Photofrin mediated PDT is performed as follows: i) Photofrin is administrated via intravenous injection with 24-48 hours incubation, ii) activated Photofrin with light at wavelength 630 nm. It has had promising results in controlling recurrence in breast cancer, brain tumors, and head and neck neoplasms [91-93]. However, drawbacks of Photofrin such as low singlet oxygen quantum yield at 630 nm and long clearance time limited the application of Photofrin in cancer treatment. The drug is ineffective to generate singlet oxygen at 630 nm activation and require long time for treatment. A 24-48 hours drug incubation time favors concentration of Photofrin in rapid proliferating tissue such as tumor and to maximize the difference of Photofrin concentration in normal tissue and rapid proliferating tissue. Slow clearance time is another drawback of Photofrin mediated PDT. HpD remains in tissue for 4-6 weeks after injection and thus patients are advised to protect themselves from exposure to sunlight or bright light for a 4-8 weeks period in order to avoid skin photosensitization [94,95].
5-aminolevulinic Acid Hexyl Ester Derivatives and Protoporphyrin (PpIX)
A number of 2nd generation photosensitizers of different chemical
families were synthesized in the late 1980s to offer potential advantages
over the 1st generation photosensitizers, including higher chemical
purity, better tumour selectivity and faster clearance [86,96].
5-aminolevulinic acid (5-ALA), the precurous substance
of protoporphyrin, is one of the FDA approved 2nd generation
photosensitizers and is popularly used for in vivo and in vitro studies
over the past decade. It has been applied on cancers such as skin
cancer, gastrointestinal adenocarcinoma, Bowen disease and basal
cell carcinoma and promising result was obtained [97,98].
5-ALA is the precursor (also known as pro-drug) of an
endogenous photosensitizer, protoporphyrin IX (PpIX), involved
in the heme bio-synthesis pathway. 5-ALA accumulated in
mitochondria region will be converted into PpIX through oxidation
(Figure 1) and will further convert into iron (II) protoporphyrin
(protoheme) by ferrochelatase in the present of iron. By adding excess
exogenous 5-ALA, more PpIX will be generated and accumulated in
the cells as the rate of ALA transform into PpIX is greater than the
rate of PpIX convert into protoheme. PpIX is temporary accumulated
in cells with high exogenous ALA, thus within tumour cells (Figure
1) [97]. Cells will become cytotoxic when PpIX is photo-activated
with specific wavelength. The wavelength applied depends on the
absorption wavelength of photosensitizer. Low wavelength will have
a poor penetration rather than high wavelength. However, of the
light applied has a too high wavelength may causing photo-damage
to the surface cells. Thus the optimum wavelength applied is always
between 620-650 nm [97,99]. There are several factors affecting the
PpIX accumulation in tumour cells, such as the uptake of 5-ALA,
concentration of iron and activity of the enzyme ferrothelatase [100].
The major side effect of 5-ALA refer to its poor penetration
ability via the biological membrane. The poor biological membrane
penetration of 5-ALA was due to its hydrophilic properties [97].
Therefore, a more effective and powerful derivatives of 5-ALA was
developed and is known as the 5-ALA hexyl ester (H-ALA). H-ALAis
one of the 5-ALA derivatives with increase in lipophilic properties
by adding a long lipophilic chain (hexyl group) to 5-ALA. The hexyl
group added to 5-ALA result as better penetration of 5-ALA into
cytoplasm. Study shows that 60 fold increase of ALA is needed to
produce same amount of PpIX accumulation inside cells compare to
H-ALA [101-109].
Figure 1
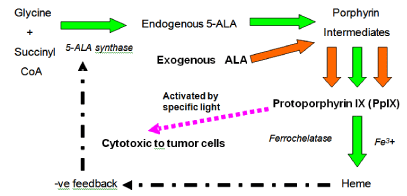
Figure 1
Effect of exogenous 5-ALA in the pathway of heme formation
by endogenous ALA.
Increase of exogenous ALA results in higher PpIX formation and
accumulation, as insufficient ferrochelatase for the conversion of PpIX to
heme. Appropriate light irradiation activates the accumulated PpIX resulting
in tumour cell killing.
Foscan® and FosPeg® (Biolitec AG)-Mesotetrahydroxyphenylchlorin (mTHPC) and Its Pegylated Liposomes Form
Meta-tetra (hydroxyphenyl) chlorine (mTHPC) is another 2nd
generation photosensitizers with hyrophobic nature that has excellent
photocytotoxicity. It is a clinically approved photosensitizer in USA,
Europe and UK and has been shown to be highly effective in treating
disease like basal cell carcinoma, prostate and pancreatic cancer [110-
113]. The typical mTHPC mediated PDT is performed as follow: i)
mTHPC is administrated via intravenous injection, ii) a 24-96h drug
incubation followed by the light activation at 652 nm wavelength. The
mTHPC, a chlorine-like photosensitizer contains active ingredient
temoporfin and is derived from the reduction of porphyrins. Bonnet
was the one who first synthesized temoporfin as pure compound
by reduction reaction and reported its photophysical properties
and photo-cytotoxicity in 1989 [114]. It has a hydrophobic nature
which ensures rapid penetration across biological membrane and
localization at critical intracellular membranous organelles [115]. The
absorption peak of mTHPC shifts to the longer wavelength of 652 nm
in the red spectrum, which favours for a deeper tissue penetration. It
showed a 10 times higher extinction coefficient than that of Photofrin
and results as a shorter incubation time [116]. Therefore, it is a
more potent photosensitizer (approximated 100 times greater) than
Photofrin or 5-ALA [95,117-119].
However, the major drawbacks of mTHPC are the bio-distribution,
clearance and selectivity of tumour uptake. These problems are related
to the photochemical properties of mTHPC. The hydrophobicity
leads to poor solubility of mTHPC in physiologically acceptable
media, which complicated its formulation and administration. It is
soluble in inorganic solvent but is insoluble in all aqueous media.
Another side effect of mTHPC is the bio-distrubution. Hydrophobic
mTHPC forms aggregates, which decreases in photo-activity and
binds strongly to serum protein. mTHPC will also accumulates in
subcutaneous fat tissue near intravenous administration and prolongs
the clearance time to 4-6 weeks after injection. All these problem of
mTHPC urge the development of drug delivery system – liposomal
formulation of mTHPC [120,121].
Current development of photosensitizer, also known as the 3rd
generation of photosensitizers, amis at improve the drug delivery
approached, such as biological modifications like antibody conjugate
or liposome conjugate [77,84,122-124]. The aim of using liposomes
for carrier and delivery systems is to improve its therapeutic
effects by solubilize the photosensitizer at suitable concentration,
increase drug uptake as well as tumour eradication [125-128]. The
formation of mTHPC contained in pegylated liposomes is known
as FosPeg® (Biolitec AG). The lipocompatible polymer Polyethylene
glycol (PEG) has been chemically linked to the outer surface of the
liposome. They function as i) stabilizer to stabilise the liposome, ii) increases its hydrophilicity to minimise the binding of liposomes to
opsonins (minimize the loss of liposome from circulation), iii) inhibit
the release of hydrophobic PSs to the liposomal membrane system via
binding with the serum protein, iv) avoid recognition by the host’s
immune system [120,129-131]. Modification of liposomes with longcirculating
poly ethylene glycol (PEG) could improve the bioavility of
m-THPC and could improve the therapeutic index of encapsulated
drugs [115,120,125-127,132,133].
Light Sources and Light Dose for PDT
Development of new light source is one of the important factors
to improve the efficiency of PDT. Light sources commonly used for
PDT including laser, laser diodes, light-emitting diodes and filtered
board-band light. Three main criteria must be fulfilled for an ideal
light source, including i) light spectrum emitted must correspond
to the absorption spectrum of the photosensitizer selected, ii) the
wavelength must be long enough in order to achieve a deep tissue
penetration, iii) sufficient photon energy should be provided to
maximize the quantum yield of singlet oxygen [134-136]. The choice
of light source depends on the photosensitizer and the depth of tissue
need to be penetrated. In early days, conventional light sources such
as halogen lamps and xenon lamps were employed for the in vitro
and in vivo PDT studies. As board-band light sources, the specific
wavelength output is achieved by adapting different filters. Usage
of these non-coherentive conventional light sources to activate
photosensitizers mainly due to the following, i) the ease of use, ii)
low cost iii) and large treatment area [124]. Incoherent and coherent
light are commonly employed for PDT and usually show similar
efficiencies [137].
Advanced light sources such as laser, laser diodes and lightemitting
diodes (LED) have become the light source of choice as they
could produce coherentive light with monochromatic wavelength
that allows easy calculation of light dosimetry. Light generated from
these light sources could be delivered down an optical fibre to the
desired site of treatment to achieve a greater selectivity [138,139].
Among these, LED based light sources become more important and
raise peoples’ concern in recent time because of the advantages such
as relatively cheap, considerably small in size and relatively mobile,
produce cool light which need not to have cooling units, and reliable
[140]. Studies of light dosimetry focus on the calculation of power
needed for a PDT treatment. The power applied for a PDT treatment
is defined as the light dose, which expressed as the energy delivered
per unit area (J/cm2). The term light dose is used to describe the
relationship between light power density and irradiation time in
order to reflect the amount of light that reaches the surface of the
tumour in a period of time. There are some criteria for selecting
an appropriate light dose, including the penetration depth to the
tissue, low light intensity and short treatment time to avoid heating
[141-143]. In parallel to the light dose, drug dose is also important
to formula the overall photodynamic dose (light dose x drug dose).
Factors affect the outcome of PDT including, photosensitizers (PSs)
applied, intracellular concentration of PSs, localization of PSs,
quantum energy obtained by the PSs and concentration of molecular
oxygen presence.
PDT Effect on NPC – Molecular Targets and Drug Resistance
The development of improved therapeutic strategies, such as PDT and immunotherapy, shed light on the development of NPC treatment [108,144-148]. Our group demonstrated promising outcomes from a number of in vitro studies concerning the PDT effect using several PSs including hypericin, mTHPC, merocyanine 540, 5-ALA and hexyl- ALA on NPC/HK1, NPC/CNE1 and NPC/CNE2 cells. We found that such photo-cytotoxicities were mediated through the mitogen activated protein kinase (MAPK) signal pathways. In response to PDT, expression of MAPK signals - ERK and p38 and Epidermal Growth Factor receptor (EGFR) signal pathways were found to be inhibited and associated with the apoptotic tumor cell death. Our studies also revealed the PDT effect on EBV viral components. PDT triggered LMP1 related mRNA and miRNAs expression, which inhibit NPC cell proliferation and cell migration. The PDT efficacy was also proved to be independent to the drug resistance properties [108,119,132, 149-158]. Other study reported the association between suppression and anti-proliferative activities induced by PDT using different cell models. Lai and his colleagues showed that PDT has an immuno-enhancing effect in NPC patients by increasing natural killer cells and interleukin-2 [159]. Another group from Hong Kong has illustrated similar outcomes by using other PSs curcumin and Zn-BC-AMon NPC/CNE2 cells and NPC/HK1 cells respectively [156,158,160]. Preliminary clinical studies using hematoporphyrin and temoporfin for the treatment of the local and recurrence of NPC after curative radiotherapy found encouraging result for residual or recurrent NPC restricted locally to the nasopharynx [161,162]. Studies also revealed that PDT could modulate the inflammatory cytokine production and angiogenic factors production [163-165].
Conclusion
PDT could be one of the best choices over the conventional cancer therapies for NPC patients, particularly for those with distance recurrence and multi drug resistance properties developed.
References
- Sizhong Z, Xiukung G, Yi Z. Cytogenetic studies on an epithelial cell line derived from poorly differentiated nasopharyngeal carcinoma. Int J Cancer. 1983; 31: 587-590.
- Vokes EE, Liebowitz DN, Weichselbaum RR. Nasopharyngeal carcinoma. Lancet. 1997; 350: 1087-1091 .
- Jia WH, Huang QH, Liao J, Ye W, Shugart YY, Liu Q, et al. Trends in incidence and mortality of nasopharyngeal carcinoma over a 20-25 year period (1978/1983-2002) in Sihui and Cangwu counties in southern China. BMC cancer. 2006; 6: 178.
- Hospital Authority: Hong Kong Cancer Registry Hong Kong. 2013.
- Yoshizaki T, Ito M, Murono S, Wakisaka N, Kondo S, Endo K. Current understanding and management of nasopharyngeal carcinoma. Auris, nasus, larynx. 2012; 39: 137-144.
- Chua ML, Wee JT, Hui EP, Chan AT. Nasopharyngeal carcinoma. Lancet. 2016; 387: 1012-1024.
- Brennan B. Nasopharyngeal carcinoma. Orphanet J Rare Dis. 2006; 1: 23.
- Tulalamba W, Janvilisri T. Nasopharyngeal carcinoma signaling pathway: an update on molecular biomarkers. Int J Cell Biol. 2012: 594681.
- Zhang L, Chen QY, Liu H, Tang LQ, Mai HQ. Emerging treatment options for nasopharyngeal carcinoma. Drug Des Devel Ther. 2013; 7: 37-52.
- Okano M, Thiele GM, Davis JR, Grierson HL, Purtilo DT. Epstein-Barr virus and human diseases: recent advances in diagnosis. Clin Microbiol Rev. 1988; 1: 300-312.
- Farrell PJ, Cludts I, Stuhler A. Epstein-Barr virus genes and cancer cells. Biomedicine & pharmacotherapy = Biomedecine & pharmacotherapie. 1997; 51: 258-267.
- Herrmann K, Niedobitek G. Epstein-Barr virus-associated carcinomas: facts and fiction. J Pathol. 2003; 19: 140-145.
- He JR, Tang LY, Yu DD, Su FX, Song EW, Lin Y, et al. Epstein-Barr virus and breast cancer: serological study in a high-incidence area of nasopharyngeal carcinoma. Cancer Lett. 2011; 309: 128-136.
- Tsao SW, Tsang CM, Pang PS, Zhang G, Chen H, Lo KW. The biology of EBV infection in human epithelial cells. Semin Cancer Biol. 2012; 22: 137-143.
- Zheng H, Li LL, Hu DS, Deng XY, Cao Y. Role of Epstein-Barr virus encoded latent membrane protein 1 in the carcinogenesis of nasopharyngeal carcinoma. Cellu Mol Immunol. 2007; 4: 185-196.
- Raab-Traub N. Epstein-Barr virus in the pathogenesis of NPC. Semin Cancer Biol. 2002 ; 12: 431-441.
- Yang CF, Yang GD, Huang TJ, Li R, Chu QQ, Xu L, et al. EB-virus latent membrane protein 1 potentiates the stemness of nasopharyngeal carcinoma via preferential activation of PI3K/AKT pathway by a positive feedback loop. Oncogene. 2016; 35: 3419-3431.
- Zeng Z, Fan S, Zhang X, Li S, Zhou M, Xiong W, et al. Epstein-Barr virus-encoded small RNA 1 (EBER-1) could predict good prognosis in nasopharyngeal carcinoma. Clin transl oncol. 2016 ; 18: 206-211.
- Ahmed HG, Suliman RS, El Aziz MS, Alshammari FD. Molecular screening for Epstein Barr virus (EBV) among Sudanese patients with nasopharyngeal carcinoma (NPC). Infect agent cancer. 2015; 10: 6.
- Raab-Traub N. Epstein–Barr virus in the pathogenesis of NPC. semin in Cancer Biol. 2002 ;12: 431-441.
- Goormachtigh G, Ouk TS, Mougel A, Tranchand-Bunel D, Masy E, Le Clorennec C, et al. Autoactivation of the Epstein-Barr virus oncogenic protein LMP1 during type II latency through opposite roles of the NF-kappaB and JNK signaling pathways. J virol. 2006 ;8: 7382-7393.
- Kung CP, Raab-Traub N. Epstein-Barr virus latent membrane protein 1 induces expression of the epidermal growth factor receptor through effects on Bcl-3 and STAT3. Journal of virology. 2008; 82: 5486-5493.
- Dawson CW, Port RJ, Young LS. The role of the EBV-encoded latent membrane proteins LMP1 and LMP2 in the pathogenesis of nasopharyngeal carcinoma (NPC). Semin Cancer Biol. 2012; 22: 144-153.
- Teramoto N, Maeda A, Kobayashi K, Hayashi K, Oka T, Takahashi K, et al. Epstein-Barr virus infection to Epstein-Barr virus-negative nasopharyngeal carcinoma cell line TW03 enhances its tumorigenicity. Lab invest. 2000; 80: 303-312 .
- Eliopoulos AG, Young LS. LMP1 structure and signal transduction. Semin Cancer Biol. 2001; 11: 435-444.
- Yoshizaki T, Sato H, Murono S, Pagano JS, Furukawa M. Matrix metalloproteinase 9 is induced by the Epstein-Barr virus BZLF1 transactivator. Clin Exp Metastasis. 1999; 17: 431-436.
- He ML, Luo MX, Lin MC, Kung HF. MicroRNAs: potential diagnostic markers and therapeutic targets for EBV-associated nasopharyngeal carcinoma. Biochim Biophys Acta. 2012 ; 182: 1-10.
- Chan AT. Nasopharyngeal carcinoma. Ann Oncol. 2010; 21: vii308-312.
- Lo AK, To KF, Lo KW, Lung RW, Hui JW, Liao G, et al. Modulation of LMP1 protein expression by EBV-encoded microRNAs. Proc Natl Acad Sci USA. 2007; 104: 16164-16169.
- Tsai CL, Li HP, Lu YJ, Hsueh C, Liang Y, Chen CL, et al. Activation of DNA methyltransferase 1 by EBV LMP1 Involves c-Jun NH(2)-terminal kinase signaling. Cancer res. 2006 ; 6: 11668-11676.
- Smith PR, de Jesus O, Turner D, Hollyoake M, Karstegl CE, Griffin BE, et al. Structure and coding content of CST (BART) family RNAs of Epstein-Barr virus. J virol. 2000; 74: 3082-3092.
- Smith P. Epstein-Barr virus complementary strand transcripts (CSTs/BARTs) and cancer. Semin Cancer Biol. 2001; 11: 469-476.
- Takada K. Role of EBER and BARF1 in nasopharyngeal carcinoma (NPC) tumorigenesis. Semin Cancer Biol. 2012; 22: 162-165.
- Marquitz AR, Raab-Traub N. The role of miRNAs and EBV BARTs in NPC. Semin Cancer Biol. 2012; 22: 166-172.
- Chen SJ, Chen GH, Chen YH, Liu CY, Chang KP, Chang YS, et al. Characterization of Epstein-Barr virus miRNAome in nasopharyngeal carcinoma by deep sequencing. PloS one. 2010; 5.
- Cosmopoulos K, Pegtel M, Hawkins J, Moffett H, Novina C, Middeldorp J, et al. Comprehensive profiling of Epstein-Barr virus microRNAs in nasopharyngeal carcinoma. J virol. 2009; 8: 2357-2367.
- Zhao Y, Wang Y, Zeng S, Hu X. LMP1 expression is positively associated with metastasis of nasopharyngeal carcinoma: evidence from a meta-analysis. J clin pathol. 2012; 65: 41-45.
- Choy EY, Siu KL, Kok KH, Lung RW, Tsang CM, To KF, et al. An Epstein-Barr virus-encoded microRNA targets PUMA to promote host cell survival. The J Exp Med. 2008; 205: 2551-2560.
- Iizasa H, Wulff BE, Alla NR, Maragkakis M, Megraw M, Hatzigeorgiou A, et al. Editing of Epstein-Barr virus-encoded BART6 microRNAs controls their dicer targeting and consequently affects viral latency. J biol chem. 2010 ; 285: 33358-33370.
- Barth S, Pfuhl T, Mamiani A, Ehses C, Roemer K, Kremmer E, et al. Epstein-Barr virus-encoded microRNA miR-BART2 down-regulates the viral DNA polymerase BALF5. Nucleic Acids Res. 2008 ; 36: 666-675.
- Lo AK, Dawson CW, Jin DY, Lo KW. The pathological roles of BART miRNAs in nasopharyngeal carcinoma. J pathol. 2012 ; 227: 392-403.
- Gourzones C, Jimenez AS, Busson P. Profiling of Epstein-Barr virus-encoded microRNAs in nasopharyngeal carcinoma reveals potential biomarkers and oncomirs. Cancer. 2012 ; 118: 4634.
- Wong AMG, Tsang JW,Kong KL,Kwong DL, Guan X-Y. Reply to profiling of Epstein-Barr virus-encoded microRNAs in nasopharyngeal carcinoma reveals potential biomarkers and oncomirs. Cancer. 2012; 118: 698-710.
- Riley KJ, Rabinowitz GS, Yario TA, Luna JM, Darnell RB, Steitz JA. EBV and human microRNAs co-target oncogenic and apoptotic viral and human genes during latency. EMBO J. 2012 ; 31: 2207-2221.
- Li G, Wu Z, Peng Y, Liu X, Lu J, Wang L, et al. MicroRNA-10b induced by Epstein-Barr virus-encoded latent membrane protein-1 promotes the metastasis of human nasopharyngeal carcinoma cells. Cancer letters. 2010; 299: 29-36.
- Capella MA, Capella LS. A light in multidrug resistance: photodynamic treatment of multidrug-resistant tumors. J Biomed sci. 2003; 10: 361-366.
- Stavrovskaya AA. Cellular mechanisms of multidrug resistance of tumor cells. Biochemistry (Mosc). 2000 ; 65: 95-106.
- Szakacs G, Paterson JK, Ludwig JA, Booth-Genthe C, Gottesman MM. Targeting multidrug resistance in cancer. Nat Rev Drug Discov. 2006; 5: 219-34.
- Wang XK, Fu LW. Interaction of tyrosine kinase inhibitors with the MDR- related ABC transporter proteins. Curr Drug metab. 2010; 11: 618-628.
- Teodori E, Dei S, Martelli C, Scapecchi S, Gualtieri F. The functions and structure of ABC transporters: implications for the design of new inhibitors of Pgp and MRP1 to control multidrug resistance (MDR). Curr Drug Targets. 2006 ; 7: 893-909.
- Aszalos A. Drug-drug interactions affected by the transporter protein, P-glycoprotein (ABCB1, MDR1) I. Preclinical aspects. Drug Discov Today. 2007 ; 12: 833-837.
- Solazzo M, Fantappie O, Lasagna N, Sassoli C, Nosi D, Mazzanti R. P-gp localization in mitochondria and its functional characterization in multiple drug-resistant cell lines. Exp Cell Res. 2006 ; 312: 4070-4078.
- Chen CL, Sheen TS, Lou IU, Huang AC. Expression of multidrug resistance 1 and glutathione-S-transferase-Pi protein in nasopharyngeal carcinoma. Hum Pathol. 2001 ; 32: 1240-1244.
- Larbcharoensub N, Leopairat J, Sirachainan E, Narkwong L, Bhongmakapat T, Rasmeepaisarn K, et al. Association between multidrug resistance-associated protein 1 and poor prognosis in patients with nasopharyngeal carcinoma treated with radiotherapy and concurrent chemotherapy. Hum Pathol. 2008; 39: 837-845.
- Tsuzuki H, Fujieda S, Sunaga H, Sugimoto C, Tanaka N, Saito H. Expression of multidrug resistance-associated protein (MRP) in head and neck squamous cell carcinoma. Cancer Lett. 1998; 126: 89-95.
- Farber S, Diamond LK, Robert D. Mercer M.D, Robert F. Sylvester,jr, James A. Wolff. Temporary remissions in acute leukemia in children produced by folic acid antagonist, 4-aminopteroyl-glutamic acid. N Engl J M. 1948; 238: 787-793.
- Juliano RL, Ling V. A surface glycoprotein modulating drug permeability in Chinese hamster ovary cell mutants. Biochimic Biophysic Acta. 1976; 455: 152-162.
- Leslie EM, Deeley RG, Cole SP. Multidrug resistance proteins: role of P-glycoprotein, MRP1, MRP2, and BCRP (ABCG2) in tissue defense. Toxicol Appl Pharmacol. 2005 ; 204: 216-237.
- Sauna ZE, Kim IW, Ambudkar SV. Genomics and the mechanism of P-glycoprotein (ABCB1). Journal of bioenergetics and biomembranes. 2007 ; 39: 481-487.
- Rosenberg MF, Callaghan R, Ford RC, Higgins CF. Structure of the multidrug resistance P-glycoprotein to 2.5 nm resolution determined by electron microscopy and image analysis. J Biol Chem. 1997; 272: 10685-10694.
- Ambudkar SV, Lelong IH, Zhang J, Cardarelli CO, Gottesman MM, Pastan I. Partial purification and reconstitution of the human multidrug-resistance pump: characterization of the drug-stimulatable ATP hydrolysis. Pro Natl Acad Sci USA. 1992; 89: 8472-8476.
- Wang EJ, Casciano CN, Clement RP, Johnson WW. Two transport binding sites of P-glycoprotein are unequal yet contingent: initial rate kinetic analysis by ATP hydrolysis demonstrates intersite dependence. Biochim Biophys Acta. 2000; 1481: 63-74.
- Ramachandra M, Ambudkar SV, Chen D, Hrycyna CA, Dey S, Gottesman MM, et al. Human P-glycoprotein exhibits reduced affinity for substrates during a catalytic transition state. Biochemistry. 1998 ; 37: 5010-5019.
- Senior AE, Bhagat S. P-glycoprotein shows strong catalytic cooperativity between the two nucleotide sites. Biochemistry. 1998; 37: 831-836.
- Sauna ZE, Ambudkar SV. Evidence for a requirement for ATP hydrolysis at two distinct steps during a single turnover of the catalytic cycle of human P-glycoprotein. Proc Natl Acad Sci U S A. 2000; 97: 2515-2520.
- Sauna ZE, Ambudkar SV. Characterization of the catalytic cycle of ATP hydrolysis by human P-glycoprotein. The two ATP hydrolysis events in a single catalytic cycle are kinetically similar but affect different functional outcomes. J biol chem. 2001; 27: 11653-11661.
- Tao Q, Chan AT. Nasopharyngeal carcinoma: molecular pathogenesis and therapeutic developments. Expert Rev Mol Med. 2007 ; 9: 1-24.
- Chen J, Dassarath M, Yin Z, Liu H, Yang K, Wu G. Radiation induced temporal lobe necrosis in patients with nasopharyngeal carcinoma: a review of new avenues in its management. Radiat Oncol. 2011; 6:128.
- Suarez C, Rodrigo JP, Rinaldo A, Langendijk JA, Shaha AR, Ferlito A. Current treatment options for recurrent nasopharyngeal cancer. Eur Arch otorhinolaryngol. 2010; 267: 1811-1824.
- Agostinis P, Berg K, Cengel KA, Foster TH, Girotti AW, Gollnick SO, et al. Photodynamic therapy of cancer: an update. CA Cancer J Clin. 2011; 61: 250-281.
- Bredell MG, Besic E, Maake C, Walt H. The application and challenges of clinical PD-PDT in the head and neck region: a short review.J photochem photobiol B. 2010; 10: 185-190.
- Brown SB, Brown EA, Walker I. The present and future role of photodynamic therapy in cancer treatment.lancet oncol. 2004; 5: 497-508.
- Plaetzer K, Krammer B, Berlanda J, Berr F, Kiesslich T. Photophysics and photochemistry of photodynamic therapy: fundamental aspects. Lasers Med sci. 2009; 2: 259-268.
- Robertson CA, Evans DH, Abrahamse H. Photodynamic therapy (PDT): a short review on cellular mechanisms and cancer research applications for PDT. J photochem photobiol B. 2009; 96: 1-8.
- Dolmans DE, Fukumura D, Jain RK. Photodynamic therapy for cancer. Nat Rev Cancer. 2003; 3: 380-387.
- Juzeniene A, Moan J. The history of PDT in Norway Part one: Identification of basic mechanisms of general PDT. Photodiagnosis Photodyn Ther. 2007; 4: 3-11.
- Calzavara-Pinton PG, Venturini M, Sala R. Photodynamic therapy: update 2006. Part 1: Photochemistry and photobiology. J Eur Acad Dermatology and Venereol. 2007; 21: 293-302.
- Huang Z, Xu H, Meyers AD, Musani AI, Wang L, Tagg R, et al. Photodynamic therapy for treatment of solid tumors--potential and technical challenges. Technol cancer Res Treat. 2008; 7: 309-320.
- Rumie Vittar NB, Lamberti MJ, Pansa MF, Vera RE, Rodriguez ME, Cogno IS, et al. Ecological photodynamic therapy: new trend to disrupt the intricate networks within tumor ecosystem. Biochim Biophys Acta. 2013; 1835: 86-99.
- Kolarova H, Nevrelova P, Tomankova K, Kolar P, Bajgar R, Mosinger J. Production of reactive oxygen species after photodynamic therapy by porphyrin sensitizers. Gen physiol biophys. 2008; 27: 101-105.
- Juarranz A, Jaen P, Sanz-Rodriguez F, Cuevas J, Gonzalez S. Photodynamic therapy of cancer. Basic principles and applications. Clin transl oncol 2008; 10: 148-154.
- Qiang YG, Yow CM, Huang Z. Combination of photodynamic therapy and immunomodulation: current status and future trends. Med Res Rev. 2008; 28: 632-644.
- Juarranz A, Jaen P, Rodriguez FS, Cuevas J, Gonzailez S. Photodynamic therapy of cancer: Basic principles and applications. Clin Transl Oncol. 2008; 10: 148-154.
- Zamadar M, Ghosh G, Mahendran A, Minnis M, Kruft BI, Ghogare A, et al. Photosensitizer drug delivery via an optical fiber. J Am Chem Soc. 2011; 133: 7882-7891.
- Garland MJ, Cassidy CM, Woolfson D, Donnelly RF. Designing photosensitizers for photodynamic therapy: strategies, challenges and promising developments. Future Med Chem. 2009; 1: 667-691.
- Dougherty TJ, Gomer CJ, Henderson BW, Jori G, Kessel D, Korbelik M, et al. Photodynamic therapy. J Natl Cancer Inst. 1998; 90: 889-905.
- Dougherty TJ. Photosensitization of malignant tumors. Semin surg oncol. 1986; 2: 24-37.
- Dougherty TJ. Studies on the structure of porphyrins contained in Photofrin II. Photochemistry and photobiology. 1987; 46: 569-573.
- Yang PW, Hung MC, Hsieh CY, Tung EC, Wang YH, Tsai JC, et al. The effects of Photofrin-mediated photodynamic therapy on the modulation of EGFR in esophageal squamous cell carcinoma cells. Lasers Med Sci. 2013; 28: 605-614.
- Dougherty TJ. Hematoporphyrin as a photosensitizer of tumors. Photochemistry and photobiology. 1983; 38: 377-379.
- Cengel KA, Glatstein E, Hahn SM. Intraperitoneal photodynamic therapy. Cancer Treat Res. 2007; 134: 493-514.
- Hsieh YJ, Wu CC, Chang CJ, Yu JS. Subcellular localization of Photofrin determines the death phenotype of human epidermoid carcinoma A431 cells triggered by photodynamic therapy: when plasma membranes are the main targets. J cell physiol. 2003; 194: 363-375.
- Leunig A, Staub F, Peters K, Heimann A, Csapo C, Kempski P, et al. Relation of early photofrin uptake to photodynamically induced phototoxicity and changes of cell volume in different cell lines. European Journal of Cancer. 1994; 30: 6.
- Allison RR, Sibata CH. Oncologic photodynamic therapy photosensitizers: a clinical review. Photodiagnosis Photodyn Ther. 2010; 7: 61-75.
- Yano S, Hirohara S, Obata M, Hagiya Y, Ogura S-i, Ikeda A, et al. Current states and future views in photodynamic therapy. Journal of Photochemistry and Photobiology C: Photochemistry Reviews. 2011; 12: 22.
- Pushpan SK, Venkatraman S, Anand VG, Sankar J, Parmeswaran D, Ganesan S, et al. Porphyrins in photodynamic therapy - a search for ideal photosensitizers. Curr Med Chem Anticancer Agents. 2002; 2: 187-207.
- Peng Q, Warloe T, Berg K, Moan J, Kongshaug M, Giercksky KE, et al. 5-Aminolevulinic acid-based photodynamic therapy. Clinical research and future challenges. Cancer. 1997; 79: 2282-2308.
- Morton CA, Whitehurst C, McColl JH, Moore JV, MacKie RM. Photodynamic therapy for large or multiple patches of Bowen disease and basal cell carcinoma. Arch Dermatol. 2001; 137: 319-324.
- Kennedy JC, Marcus SL, Pottier RH. Photodynamic therapy (PDT) and photodiagnosis (PD) using endogenous photosensitization induced by 5-aminolevulinic acid (ALA): mechanisms and clinical results. J clin laser Med Surg. 1996; 14: 289-304.
- Brunner H, Hausmann F, Knuechel R. New 5-aminolevulinic acid esters--efficient protoporphyrin precursors for photodetection and photodynamic therapy. Photochem photobiol. 2003; 78: 481-486.
- Eleouet S, Rousset N, Carre J, Bourre L, Vonarx V, Lajat Y, et al. In vitro fluorescence, toxicity and phototoxicity induced by delta-aminolevulinic acid (ALA) or ALA-esters. Photochem Photobiol. 2000; 71: 447-454.
- Casas A, Perotti C, Saccoliti M, Sacca P, Fukuda H, Batlle AdC. ALA and ALA hexyl ester in free and liposomal formulations for the photosensitisation of tumor organ cultures. Br J cancer. 2002; 86: 7.
- Juzeniene A, Ma LW, Juzenas P, Iani V, Lange N, Moan J. Production of protoporphyrin IX from 5-aminolevulinic acid and two of its esters in cells in vitro and tissues in vivo. Cell Mol Biol. 2002; 48: 911-916.
- Dognitz N, Salomon D, Zellweger M, Ballini JP, Gabrecht T, Lange N, et al. Comparison of ALA- and ALA hexyl-ester-induced PpIX depth distribution in human skin carcinoma. J Photochem Photobiol B. 2008; 93: 140-148.
- Bruce-Micah R, Huttenberger D, Freitag L, Cullum J, Foth HJ. Pharmacokinetic of ALA and h-ALA induced porphyrins in the models Mycobacterium phlei and Mycobacterium smegmatis. J Photochem Photobiol B. 2009; 97: 1-7.
- Morrow DI, McCarron PA, Woolfson AD, Juzenas P, Juzeniene A, Iani V, et al. Hexyl aminolaevulinate is a more effective topical photosensitiser precursor than methyl aminolaevulinate and 5-aminolaevulinic acids when applied in equimolar doses. J Pharma Sci. 2010; 99: 3486-3498.
- Wu RW, Yow CM, Wong CK, Lam YH. Photodynamic therapy (PDT) - Initiation of apoptosis via activation of stress-activated p38 MAPK and JNK signal pathway in H460 cell lines. Photodiagnosis Photodyn Ther. 2011; 8: 254-263.
- Wu RW, Chu ES, Yow CM, Chen JY. Photodynamic effects on nasopharyngeal carcinoma (NPC) cells with 5-aminolevulinic acid or its hexyl ester. Cancer Lett. 2006; 24: 112-119.
- Yow CMN, Wu RWK, Huang Z. Comparison of Aminolevulinic Acid and Its Methyl Ester Mediated Photocytotoxicity on Human Nasopharyngeal Carcinoma Cells. Journal of Innovative Optical Health Sciences. 2012; 05: 1250007.
- Melnikova VO, Bezdetnaya LN, Bour C, Festor E, Gramain MP, J.L.Merlin, et al. Subcellular localization of meta-teta(hydroxyphenyl)chlorine in human tumor cells subjected to photodynamic treatment. J Photochem Photobiol B. 1999; 49: 8.
- Moore CM, Nathan TR, Lees WR, Mosse CA, Freeman A, Emberton M, et al. Photodynamic therapy using meso tetra hydroxy phenyl chlorin (mTHPC) in early prostate cancer. Lasers Surg Med. 2006; 38: 356-363.
- Baas P, Saarnak AE, Oppelaar H, Neering H, Stewart FA. Photodynamic therapy with meta-tetrahydroxyphenylchlorin for basal cell carcinoma: a phase I/II study. Br J Dermatol. 2001; 145: 75-78.
- Bown SG, Rogowska AZ, Whitelaw DE, Lees WR, Lovat LB, Ripley P, et al. Photodynamic therapy for cancer of the pancreas. Gut. 2002; 50: 9.
- Bonnett R, White RD, Winfield UJ, Berenbaum MC. Hydroporphyrins of the meso-tetra(hydroxyphenyl)porphyrin series as tumour photosensitizers. Biochem J. 1989; 261: 277-280.
- Pegaz B, Debefve E, Ballini JP, Wagnieres G, Spaniol S, Albrecht V, et al. Photothrombic activity of m-THPC-loaded liposomal formulations: pre-clinical assessment on chick chorioallantoic membrane model. Eur J Pharm Sci. 2006; 28: 134-140.
- R.Bonnett. Photodynamic therapy in historical perspective. Rev Contemp Pharmacother. 1999; 10: 17.
- Mitra S, Maugain E, Bolotine L, Guillemin F, Foster TH. Temporally and spatially heterogeneous distribution of mTHPC in a murine tumor observed by two-color confocal fluorescence imaging and spectroscopy in a whole-mount model. Photochem Photobiol. 2005; 81: 1123-1130.
- Bourre L, Rousset N, Thibaut S, Eleouet S, Lajat Y, Patrice T. PDT effects of m-THPC and ALA, phototoxicity and apoptosis. Apoptosis 2002; 7: 221-230.
- Yow CM, Chen JY, Mak NK, Cheung NH, Leung AW. Cellular uptake, subcellular localization and photodamaging effect of temoporfin (mTHPC) in nasopharyngeal carcinoma cells: comparison with hematoporphyrin derivative. Cancer lette. 2000; 157: 123-131.
- Bovis MJ, Woodhams JH, Loizidou M, Scheglmann D, Bown SG, Macrobert AJ. Improved in vivo delivery of m-THPC via pegylated liposomes for use in photodynamic therapy. J controll Release 2012; 157: 196-205.
- Senge MO. mTHPC--a drug on its way from second to third generation photosensitizer? Photodiagnosis and photodyn Ther. 2012; : 170-179.
- Kuntsche J, Freisleben I, Steiniger F, Fahr A. Temoporfin-loaded liposomes: physicochemical characterization. Euro J Pharm Sci. 2010; 40: 305-315.
- Paszko E, Ehrhardt C, Senge MO, Kelleher DP, Reynolds JV. Nanodrug applications in photodynamic therapy. Photodiagnosis Photodyn Ther. 2011; 8: 14-29.
- Triesscheijn M, Ruevekamp M, Antonini N, Neering H, Stewart FA, Baas P. Optimizing meso-tetra-hydroxyphenyl-chlorin-mediated photodynamic therapy for basal cell carcinoma. Photochem Photobiol. 2006; 82: 1686-1690.
- Buchholz J, Kaser-Hotz B, Khan T, Rohrer Bley C, Melzer K, Schwendener RA, et al. Optimizing photodynamic therapy: in vivo pharmacokinetics of liposomal meta-(tetrahydroxyphenyl)chlorin in feline squamous cell carcinoma. Clin Cancer Res. 2005; 11: 7538-7544.
- D'Hallewin MA, Kochetkov D, Viry-Babel Y, Leroux A, Werkmeister E, Dumas D, et al. Photodynamic therapy with intratumoral administration of Lipid-Based mTHPC in a model of breast cancer recurrence. Lasers Surg Med. 2008; 40: 543-549.
- Lassalle HP, Dumas D, Grafe S, D'Hallewin MA, Guillemin F, Bezdetnaya L. Correlation between in vivo pharmacokinetics, intratumoral distribution and photodynamic efficiency of liposomal mTHPC. J Control Release. 2009; 134: 118-124.
- Maeda H, Wu J, Sawa T, Matsumura Y, Hori K. Tumor vascular permeability and the EPR effect in macromolecular therapeutics: a review. J Control Release. 2000; 65: 271-284.
- Dos Santos N, Allen C, Doppen AM, Anantha M, Cox KA, Gallagher RC, et al. Influence of poly(ethylene glycol) grafting density and polymer length on liposomes: relating plasma circulation lifetimes to protein binding. Biochim Biophys Acta. 2007; 1768: 1367-1377.
- Klibanov AL, Maruyama K, Torchilin VP, Huang L. Amphipathic polyethyleneglycols effectively prolong the circulation time of liposomes. FEBS letters. 1990; 268: 235-237.
- Satomi T, Nagasaki Y, Kobayashi H, Tateishi T, Kataoka K, Otsuka H. Physicochemical characterization of densely packed poly(ethylene glycol) layer for minimizing nonspecific protein adsorption. J Nanosci Nanotechno. 2007; 7: 2394-2399.
- Wu RW, Chu ES, Huang Z, Xu CS, Ip CW, Yow CM. Effect of FosPeg(R) mediated photoactivation on P-gp/ABCB1 protein expression in human nasopharyngeal carcinoma cells. J Photochem Photobiol B. 2015; 148: 82-87.
- Wu RW, Chu ES, Huang Z, Xu CS, Ip CW, Yow CM. FosPeg(R) PDT alters the EBV miRNAs and LMP1 protein expression in EBV positive nasopharyngeal carcinoma cells. Journal of photochemistry and photobiology B, Biology. 2013; 127: 114-122.
- Nestor M, Gold M, Kauvar A, Taub A, Geronemus R, Ritvo E, et al. The use of photodynamic therapy in dermatology: results of a consensus conference. J Drugs Dermatol. 2006; 5: 15.
- Salva KA., or Photodynamic therapy: unapproved uses, dosages indications. Clin Dermatol. 2002; 20: 571-581.
- Ceburkov O, Gollnick H. Photodynamic therapy in dermatology. European journal of dermatology : EJD. 2000; 10: 568-575.
- Juzeniene A, Juzenas P, Ma LW, Iani V, Moan J. Effectiveness of different light sources for 5-aminolevulinic acid photodynamic therapy. Lasers Med Sci. 2004; 19: 139-149.
- MacCormack M. Photodynamic therapy. Adv Dermatol. 2006; 22: 40.
- Castano AP, Demidova TN, Hamblin MR. Mechanisms in photodynamic therapy: part one—photosensitizers, photochemistry and cellular localization. Photodiagnosis Photodyn Ther. 2004; 1: 279-293.
- Mitton D, Ackroyd R. A brief overview of photodynamic therapy in Europe. Photodiagnosis Photodyn Ther. 2008; 5: 103-111.
- Henderson BW, Busch TM, Snyder JW. Fluence rate as a modulator of PDT mechanisms. Lasers Surg Med. 2006; 38: 489-493.
- Kimel S, Svaasand LO, Hammer-Wilson M, Gottfried V, Cheng S, Svaasand E, et al. Demonstration of synergistic effects of hyperthermia and photodynamic therapy using the chick chorioallantoic membrane model. Laser Surg Med. 1992; 12: 432-440.
- Hopkinson HJ, Vernon DI, Brown SB. Identification and partial characterization of an unusual distribution of the photosensitizer meta-tetrahydroxyphenyl chlorin (temoporfin) in human plasma. Photochem Photobiol. 1999; 69: 482-488.
- Lin ML, Lu YC, Chung JG, Wang SG, Lin HT, Kang SE, et al. Down-regulation of MMP-2 through the p38 MAPK-NF-kappaB-dependent pathway by aloe-emodin leads to inhibition of nasopharyngeal carcinoma cell invasion. Mol carcino. 2010; 49: 783-797.
- Chan PS, Koon HK, Wu ZG, Wong RN, Lung ML, Chang CK, et al. Role of p38 MAPKs in hypericin photodynamic therapy-induced apoptosis of nasopharyngeal carcinoma cells. Photochem Photobiol. 2009; 85: 1207-1217.
- Smith C, Tsang J, Beagley L, Chua D, Lee V, Li V, et al. Effective treatment of metastatic forms of Epstein-Barr virus-associated nasopharyngeal carcinoma with a novel adenovirus-based adoptive immunotherapy. Cancer Res. 2012; 72: 1116-1125.
- Lutzky VP, Corban M, Heslop L, Morrison LE, Crooks P, Hall DF, et al. Novel approach to the formulation of an Epstein-Barr virus antigen-based nasopharyngeal carcinoma vaccine. J Virol. 2010; 84: 407-417.
- Ma BB, Hui EP, Chan AT. Systemic approach to improving treatment outcome in nasopharyngeal carcinoma: current and future directions. Cancer Sci. 2008; 9: 1311-1318.
- Bai D, Xia X, Yow CM, Chu ES, Xu C. Hypocrellin B-encapsulated nanoparticle-mediated rev-caspase-3 gene transfection and photodynamic therapy on tumor cells. Eur J Pharmcol. 2011; 650: 496-500.
- Yow C, Mak N, Leung A, Huang Z. Induction of early apoptosis in human nasopharyngeal carcinoma cells by mTHPC-mediated photocytotoxicity. Photodiagnosis Photodyn Ther. 2009; 6: 122-127.
- Yow CM, Mak NK, Szeto S, Chen JY, Lee YL, Cheung NH, et al. Photocytotoxic and DNA damaging effect of temoporfin (mTHPC) and merocyanine 540 (MC540) on nasopharyngeal carcinoma cell. Toxicol Lett. 2000; 115: 53-61.
- Yow C, Chen J, Mak N, Cheung N, Leung A. Cellular uptake, subcellular localization and photodamaging effect of Temoporn® (mTHPC) in nasopharyngeal carcinoma cells:comparison with hematoporphyrin derivative. Cancer letters. 2000; 157: 9.
- Yow CMN, W.N.Leung. Photodynamic Therapy impede Nasopharyngeal Carcinoma metastasis: Modulation of matrix metalloproteinases, MMP-2 and MMP-9. 2003.
- Wu RW, Chu ES, Huang Z, Xu CS, Ip CW, Yow CM. FosPeg® PDT alters the EBV miRNAs and LMP1 protein expression in EBV positive nasopharyngeal carcinoma cells. J Photochem Photobiol B. 2013; 127: 114-122.
- Wu R, Huang Z, Yow C. FosPeg®-mediated photocytotoxicity suppresses cancer cell growth through down-regulation of MEK pathways. Photodiagnosis and photodynamic therapy. 2011; 8: 188.
- Koon HK, Chan PS, Wu ZG, Wong RN, Lung ML, Chang CK, et al. Role of mitogen-activated protein kinase in Zn-BC-AM PDT-induced apoptosis in nasopharyngeal carcinoma cells. Cell Biochem Func. 2010; 28: 239-248.
- Xie Y, Wei ZB, Zhang Z, Wen W, Huang GW. Effect of 5-ALA-PDT on VEGF and PCNA expression in human NPC-bearing nude mice. Oncol Rep. 2009; 22: 1365-1371.
- Koon HK, Chan PS, Wong RN, Wu ZG, Lung ML, Chang CK, et al. Targeted inhibition of the EGFR pathways enhances Zn-BC-AM PDT-induced apoptosis in well-differentiated nasopharyngeal carcinoma cells. Journal of cellular biochemistry. 2009; 108: 1356-1363.
- Lai JP, Tao ZD, Xiao JY, Zhao SP, Tian YQ. Effect of photodynamic therapy on selected laboratory values of patients with nasopharyngeal carcinoma. Ann Otol, Rhinol, Laryngol. 1997; 106: 680-682.
- Koon H, Leung AW, Yue KK, Mak NK. Photodynamic effect of curcumin on NPC/CNE2 cells. J Environ Pathol, Toxicol oncol. 2006; 25: 205-215.
- Nyst HJ, Wildeman MA, Indrasari SR, Karakullukcu B, van Veen RL, Adham M, et al. Temoporfin mediated photodynamic therapy in patients with local persistent and recurrent nasopharyngeal carcinoma after curative radiotherapy: a feasibility study. Photodiagnosis Photodyn Ther. 2012; 9: 274-281.
- Tong MC, van Hasselt CA, Woo JK. Preliminary results of photodynamic therapy for recurrent nasopharyngeal carcinoma. Eur Arch otorhinolaryngol.1996; 253: 189-192.
- Koon HK, Lo KW, Leung KN, Lung ML, Chang CC, Wong RN, et al. Photodynamic therapy-mediated modulation of inflammatory cytokine production by Epstein-Barr virus-infected nasopharyngeal carcinoma cells. Cellular Mol Immunol. 2010; 7: 323-326.
- Yee KK, Soo KC, Olivo M. Anti-angiogenic effects of Hypericin-photodynamic therapy in combination with Celebrex in the treatment of human nasopharyngeal carcinoma. Int J Mol Med. 2005; 16: 993-1002.
- Du H, Bay BH, Mahendran R, Olivo M. Hypericin-mediated photodynamic therapy elicits differential interleukin-6 response in nasopharyngeal cancer. Cancer lett. 2006; 235: 202-208.