Review Article
Review of Human Hair Follicle Biology: Dynamics of Niches and Stem Cell Regulation for Possible Therapeutic Hair Stimulation for Plastic Surgeons
Gordon H. Sasaki*
Department of Plastic Surgery, Loma Linda Medical University, USA
*Corresponding author: Gordon H. Sasaki, Department of Plastic Surgery, Loma Linda Medical University, Loma Linda, Fairmount Ave, Pasadena, CA 91105, USA
Published: 23 Dec, 2016
Cite this article as: Sasaki GH. Review of Human Hair
Follicle Biology: Dynamics of Niches
and Stem Cell Regulation for Possible
Therapeutic Hair Stimulation for Plastic
Surgeons. Clin Surg. 2016; 1: 1257.
Abstract
Embryological development of a hair follicle and its cyclical adult transformation reside in quiescent niches arranged in epidermal, mesenchymal and neruroectodermal stem cell habitats that interface with each other in a hierarchical manner by cross-talking signals throughout its morphological life cycling. These compartmental growth centers trigger through their stimulating and inhibiting molecules either the induction or retardation of architectural layers and structures for de novo or post-natal follicles. Recent evidence for extrinsic regulation by transcriptional-growth factor activity by adipocytes and platelet-rich plasma for follicle development and cycling has provided potential therapeutic avenues for follicular regeneration. This review will focus on the behavior of follicular cells that play dominant roles in the development of androgenetic alopecia and chronic telogen effluvium. It is hoped that the discussed evidence-base treatment approaches may evolve into FDAapproved therapy to address hair loss and its rejuvenation for millions of men and woman.
Introduction
Schofield [1] first proposed the term, “niches”, in hematopoietic tissues to describe specialized, discrete, and slow cycling stem cell domains in micro-environments that regulated cell identity, self-renewal and differentiation. During normal homeostasis, stem cells often leave their niches and evolve into transit-amplifying (TA) cells, which dynamically proliferate and commit to terminal differentiation [2,3]. Since then, considerable characterization of stem cell niches has been reported in invertebrate systems [4,5]. In contrast, the definition and interaction of stem cell niches in mammalian system have been less well defined because of their complexity and lack of specific markers. Despite these limitations, extensive investigations [6-8] have described the architecture and cycling of de novo and post-natal hair follicles in pigmented, albino and mutant strains in mice. Notwithstanding species-to- species differences, the study of intact human hair follicles remains particularly attractive because they represent the only mini-organs that exhibit morphological and cyclical transformations, embracing recapitulation of its embryonic development from growth (anagen), apoptotic regression (catagen), to quiescence (telogen). These events are driven by compartmentalized cell types and their signaling molecules that regulate de novo hair formation in embryonic skin and new hair growth cycling in adult skin switching from dormancy to rapid cell division during cycling. Our current incomplete understanding of the molecular controls of follicle induction and morphogenesis is almost entirely based on analysis of invertebrates and mutant mice. Therefore, caution is advised in assuming that exactly the same controls occur in human hair follicle development [9]. This review investigates recent findings on these complex putative niches and their cross-talk that reveal important new insights and strategies to understand hair follicle biology, enrichment of inductive cell populations, and development of innovative therapies for hair stimulation and treatment of associated disease states.
Stem Cell Niches of Nascent Hair Follicles
Hair follicles are organized into a number of heterogeneous cell types arranged in multiple epidermal (epithelial), dermal (mesenchymal) and neuroectodermal stem cell habitats that interface with each other in a hierarchical manner for embryonic morphogenesis and post-natal cycling [10,11]. Eventually, these quiescent stem cells proliferate and differentiate into epithelial lineages (sebocytes, eccrine cells, keratinocytes, trichocytes of the outer and inner root sheaths), mesenchymal lineages (fibroblasts of the dermal papilla and dermal sheath, arrector pili muscle), and neural crest lineages (melanocytes). Generally, mesenchymal cells are believed to play dominant roles as inducers, while epithelial cells act as responders during the process of nascent and cycling follicle formation through reciprocal and complex signaling molecules (in CAPITALS) by transcription factors(in italic) [12,13].
Epithelial Contributions to Nascent Hair Follicle Development
In a review by Schmidt-Ulrich and Paus [9], de novo initiation of
follicle development in mice embryos has been found to be contingent
upon reciprocal communications under key roles of canonical
WNT via β-catenin pathways and other regulatory activating or
inhibitory proteins (transcription factors, receptors and ligands, and
adhesion molecules) between the single-layered primitive epidermal
ectodermal cells and dermal mesenchymal cells [14,15]. It remains
unclear, however, when and how embryonal stem cell niches become
established and whether their gene expressions by transcription
factors are functionally transferred to progeny cells in stem cell niches
of the first and subsequent post-natal cycles.
Initially, undifferentiated ectodermal cells begin to gather,
elongate and organize mainly under EDaA1/EDaR/NF-кϐ and
NOGGIN/Lef-1 pathways [16,17] into microscopically identifiable
placodes which are transformed chiefly by Sonic Hedgehog (SHH)
activation of Gli2 pathways into primary hair germ centers [18]
(Figure 1). In vivo pulse-chase mice experiments [8,12] have
demonstrated that a population of quiescent histone H2B-GFP label
retaining cells (LRCs) with specified stem cell markers, such as Sox9
protein, are localized within the primary germ centers of epithelial
cell clusters early in nascent Stage 2 developing follicles. Sox9 protein
has emerged as a key regulator of stem cell identity in niche centers
that govern de novo follicle formation. Recent elegant embryonic
pulse-chase studies [9,19] established that early LRC Sox9 cells are
direct precursors of LRCs that reside in both the embryonic and adult
bulge niches (see below). Without the presence of Sox9-positive cells,
normal morphologic development of nascent follicles and cycling of
post-natal follicles not only fail to occur but impaired epidermis does
not normally recover after injury.
As epithelial columns plunge downward, regulated by a myriad of
ill-understood signaling pathways [11] (SHH/Gli2, WNT/β-catenin,
NOGGIN/Lef-1, TGFβ2) and enter into the hair peg phase of Stages
3-5, LRC Sox9 expressing cells concentrate in a developing layer and
proliferate principally under similar signaling pathways to develop
into the outer root sheath [20]. As subsequent structural layers occur
to form the lower portion of the nascent mature follicle, a trail of H2BGFP-positive
LRC cells extends towards the proximal base of hair
bulb. During Stages 6-8, progeny of LRC Sox9-derived cells [21,22]
differentiate ultimately into trichocytes and highly proliferative,
transit-amplifying matrix cells, both of which play leading roles
through to the eventual formation of the shaft and other structures
in the nascent follicle.
Another specialized epithelial Sox9 stem cell niche, called the
bulge [23,24], develops as a subjacent extension from the outer root
sheath and is located proximal to the presumptive sebaceous gland
units opposite the point of insertion of the arrector pili muscle. Mice
experiments [25] have demonstrated that bulge LRC Sox9 stem cells
develop early but are not required for embryonic follicle development
which depends critically on progeny of Sox9 cells derived from the
outer root sheath. However, progeny of LRC Sox9-derived bulge stem
cells are a requirement for formation of the sebaceous lineage and
also encompass most if not all cells in the later described secondary
hair germ [26,27], both of which promote cyclic phases of post-natal
follicles in the nude mice model [6]. At the late cyto-differentiated
Stages 6-8 of follicular maturation, a variety of transcriptional factors
has been suggested to regulate lineages that form the eventual inner
root sheath, cortex and medulla of the nascent hair shaft. Data for the
exact hierarchical cascade of signals are incomplete but implicate a
central role for the transcriptional repressor CCAAT, a displacement
protein CDP, Cutl1, and the zinc-finger transcription factor GATA3
[28-29]. In developmental waves [30], melanocytes arrive to
produce pigmentation granules in the central core of the hair shaft,
accompanied by migrating hematopoietic cells and neurotrophininduced
nerve cells which form, respectively, capillary loops and
nerve innervations located around and within the mesenchymalderived
dermal papilla. At the proximal base of the follicle, the mass
of undifferentiated matrix cells, melanocytes and cells of the proximal
outer root sheath finally congregate into the onion-shaped bulb which
partially surrounds the capillary and neural network.
Mesenchymal Contributions to Nascent Hair Follicle Development
Once epithelial placodes have formed in Stage 1, epithelial cues from placode cells are needed to induce the assemblage of mesenchymal cells, as condensates, beneath the placodes/primary hair germ centers [31] (Figure 1). Dermal mesenchymal cells in their niches play pivotal roles in de novo and cycling hair formation through release of their own first molecular message of signals and transcription factors for support of proper formation of placodes [11,16,25] and induction of epithelial downward growth during early Stage 2 by fibroblast growth factors (FGF) and bone morphogenetic proteins (BMP) inhibitor (NOGGIN) [32,33]. The complex and bewildering composition and actions of the epithelial and dermal cues are being determined and may involve certain members of the WNT family, SHH/Gli2, and PDGF-A (platelet derived growth factor-A) [34-36]. Through initiating cross-talking molecular signals between these two compartments, mesenchymal cells in the dermal condensates begin their journey downward during Stages 3-5, external to the advancing epithelial columns, and eventually form into the dermal sheath and proximal dermal papilla at the base of the nascent follicle [15]. The dermal sheath composed of three collagen layers with specialized fibroblasts of mesenchymal origin [37], lies outside of the external root sheath from the bulge level downward and becomes contiguous with the base of the dermal papilla. The dermal sheath cells have been considered as cellular reservoirs for dermal papilla cells and can regenerate a new dermal papilla after its loss [38,39]. There appears to be a two-way cellular talk and cellular migration between the dermal papilla and dermal sheath whose exact signaling mechanisms (Pl3K-Akt pathway) awaits further elucidation [40]. The dermal papilla stem cells eventually reside in a complex and diverse cellular niche of the bulb at the base of the nascent follicle, and participate in epithelial-mesenchymal cross-talk through its unique gene signature molecules with surrounding epithelial cells of the matrix, outer root sheath, as well as mesenchymal cells of the dermal sheath, to induce de novo follicle development.
Figure 1
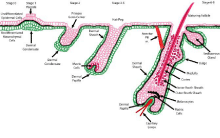
Figure 1
Schematic representation of nascent hair follicle morphogenesis during embryogenesis by specialized epithelial and mesenchymal stem cells and their
niches from Stage 0 to Stage 8.
Figure 2
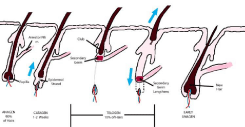
Figure 2
Schematic representation of normal mature follicle growth cycling from anagen growth, catagen regression, and telogen quiescence.
Stem Cell Niches of Post-natal Cycling of Hair Follicles
The cycling post-natal hair follicle undergoes dramatic
morphological and architectural structural transformations driven by
quiescent and proliferative niche stem cells during the anagen phase of
growth and homeostasis, transitioning to catagen phase of apoptosis
and shrinking of the lower follicle, and ending in telogen phase of
quiescent miniaturization [4] (Figure 2). The regulatory mechanisms
by which stem cells in independent sub populated compartments
become activated and mobilized are essential to follicle homeostasis.
Catagen phase highlights the important apoptotic signals from
exhausted, short-lived TA matrix progeny that initiates the start of
regressive miniaturization of the follicle. During catagen, the dermal
papilla cells provide transcriptional signals to activate the secondary
germ niche that plays a critical role during catagen’s regressive phase
in losing the lower portion of the follicle to a thin epithelial strand.
The strand is retracted upward to the base of the permanent portion
of the follicle leaving the bulge region with the small secondary germ
center which remains dormant during telogen resting phase [41].
Although the cellular relationship of the bulge and the secondary germ
remains unclear, single cell lineage tracing experiments and BrdU
labeling studies suggest, respectively, that bulge cells migrate to or
transform into the secondary germ [42] which then proliferate during
telogen to anagen transition to form the inner root sheath and hair
shaft of the new terminal hair [24,41].
After a short telogen period, a remarkable regeneration of the
miniaturized follicle is contributed from three stem cell niches
(secondary germ, bulge, and dermal papilla) to recapitulate an entire
new lower follicle and hair shaft. Investigators [7] believe that the
signaling mechanisms involved in the renewing cyclic phases mimic
those implicated during the embryonic morphogenic stages of
development. Toward the end of telogen, secondary germ cells burst
with activity fueling the release two key-step signaling pathways initiate
cycling from telogen to anagen: stabilization of β-catenin for WNT
signaling via transcriptional cofactor for Lefl;Tcf proteins [41,43];
bone morphogenetic protein (BMP) positive or negative signaling,
respectively, for induction of dermal papilla and bulge cell quiescence
[44] or initiation of morphologic and cyclic stimulation [45]. During
the brief surge of secondary germ activities, the bulge transcriptional
output remain relatively stable favoring cycle-inhibitors and
signaling repressors. Following the onset of early anagen, however,
bulge cells then unleash transcriptional and proliferative signals by
elevated levels of β-catenin [46] and are employed more sparingly to
stimulate outer root sheath, matrix cells, and inner root sheath for the
new follicle throughout anagen [47]. In summary, the secondary germ
cell niche is believed to fuel primarily the initial stages of telogenanagen
regeneration and is the first responders to dermal papilla
signals, while the bulge niche acts more as the engine that maintains
progression of the hair cycle and is the reservoir of long-term stem
cells [48]. The molecular mechanisms regulating cross-talk between
secondary germ cells and mesenchymal dermal papilla cells remain
unclear during telogen-anagen regeneration but most likely involve
the activities of BMP inhibitors, FGF (fibroblast growth factors) and
WNT effectors via β-catenin stimulation [41].
Extrinsic Regulation from Adipose Tissue for Follicle Cycling
Skin-associated white adipose tissue collections formed postnatally and existed as “intradermal fat depots” surrounding hair follicles that were distinct from subdermal/subcutaneous adipose layers [49]. These intradermal adipocytes regenerated with faster kinetics than other adipose tissue depots and their cellular dynamics parallels with the hair cycle, suggesting an inter play existed between follicles and adipocytes [50]. During cycling morphogenesis, for example, intra dermal adipocytes expanded either by cell hypertrophy0 or hyperplastic adipogenesis, doubling the skin’s thickness [51] as the follicle entered into anagen growth phase and then receded during telogen [52]. Recent studies [53,54] suggested that intradermal adipose depots increased primarily by adipogenesis, requiring the upregulation and transcriptional activity of the nuclear receptor, PPAR, in preadipocytes. Through chase BrdU pulsed mice studies [55], additional data demonstrated that preadipocyte proliferation was low during early to mid-catagen, but increased during late catagen to generate a sufficient population of mature adipocytes to induce a four-fold surge during anagen-induction. Preadipocyte numbers returned to baseline levels during homeostasis of the matured anagen phase. Taken together, these investigations indicated that there exist exquisite, but incompletely understood, regulatory mechanisms that control the stimulation or inhibition pathways for hair cycling via signals from preadipocyte precursor cells and adipocytes. To clarify the signaling importance of adipocyte-follicle interactions, a genetic mice model, lacking Early β cell factor 1 (Ebfl-/-) expression in intradermal adipocytes, sebaceous glands and dermal papilla of anagen follicles, has been shown to have less adiposederived stem cells and, therefore, reduced intradermal mature adipocytes [56]. In these mutant mice, miniaturized follicles failed to cycle into anagen and lingered instead in late catagen-telogen, suggesting the requirement of adipose-derived stem cell transition to mature adipocytes for anagen induction through the activation of the bulge stem cells. When these defective mutant mice are transplanted with WNT/β catenin expressive preadipocytes, anagen induction occurs implying need for specific adipocyte precursor lines [55]. The complicated cellular regulation for signals and transcription factors during proliferation and differentiation of adipose tissue was updated in a recent review in plastic surgery [57]. Mature intradermal adipocytes and the dermal papilla also contribute other signaling and transcription molecules that regulate bulge stem cells during the cycling process. Recent studies [58,59] have shown that mature intradermal adipocytes periodically released bone morphogenetic protein (BMP2) mRNA, an inhibitory signal, that suppressed bulge cell activity during quiescent cycling phases in early telogen and late anagen. Once BMP2 signals are extinguished, follicles became responsive to WNT/β-catenin activation signals, enabling them to complete telogen and re-enter anagen. The competing balances between BMP and WNT signals appeared to synchronize transitions from quiescence to proliferation.
Androgenetic Alopecia (AGA)
Androgenetic alopecia (AGA), commonly known as male and
female-patterned loss of hair, affects more than half of men and
women populations in the United States over age 50 [60-61] with
significant psychosocial effects in both sexes [62-63]. Although the
significant impact of androgen presence and genetic propensity are
mandatory for male AGA, much more has to be elucidated. Male AGA
is believed to be due to a polygenic mode of inheritance and presence
of dihydrotestosterone (DHT) [64]. Several lines of circumstantial
evidence demonstrated supportive roles of androgens in male AGA: 1) the absence of AGA in eunuchs and pseudo hermaphrodites [65],
lacking in functional androgen receptors (AR) and 5α-reductase
enzyme, respectively, 2) castrated post pubertal men terminating
progression of AGA [66], 3) elevated levels of 5α-reductase, DHT,
and AR in balding male scalp [67], and 4) mitigation of hair loss
with finasteride by inhibiting type II 5α-reductase activity [68]. Two
isoenzymes for 5α-reductase type I (5αR1) and type II (5αR2) are
believed to mediate the conversion of testosterone to DHT. 5αR1
is concentrated to sebaceous glands and linked in the exacerbation
of acne vulgaris, while 5αR2 has been recovered in higher levels in
follicles with male AGA [69].
Although the testosterone-receptor mechanism is not well
understood, DHT has been suggested to activate epidermal AGA
keratinocytes in the production of caspase-1 and IL-1β, regulated
at the inflammasome, immune-transcriptional levels, to facilitate
follicular miniaturization and apoptosis with each successive cycle
[70,71]. Recent studies supported these findings by demonstrating that
1) testosterone reduced TLR4 expression of immune-inflammatory
mechanisms in monocytes and macrophages [72] and that 2)
caspase-1 mediated an inflammatory cell death cycle of pyroptosis
[73]. Such contributing data in male AGA suggest that underlying
androgenic mechanisms of inflammation and innate immunity may
play significant roles in the formation of smaller, thinner, and finer
apoptotic male-patternalopecia, as defined in Norwood’s classification
of hair loss [74]. Hair loss in men is progressive with recession of the
frontal hairline and hair loss over the frontal and vertex scalp regions.
Miniaturization can abruptly occur after a few hair cycles such that
the hair does not penetrate the scalp surface.
Female pattern hair loss (FPHL) remains a poorly understood
complex in which a number of factors, such as heredity, inflammation,
hormonal or vascular influences, are under current investigation for
deleteriously affecting follicles [75-77]. While androgenic alopecia is
generally the most common form of hair loss in men, the involvement
of androgens in patterned or non-patterned hair thinning or loss has
not been as well established in women [78-79]. FPHL can often be
precipitated and exacerbated by drugs, acute stress, diet, hormonal
changes, weight loss and partum [80]. FPHL, described as regressive
or senescent alopecia, is characterized by progressive shortening
of the anagen phase, a lengthening of the latency period, and
miniaturization into villus-like hair [61]. Such changes lead to diffuse
loss of hair density, affecting primarily the vertex and frontal scalp,
defined by different grading systems [81,82].
Chronic telogen effluvium (CTE), on the other hand, is believed
to occur in apparently healthy women [83] and is attributed to an
asynchronization disorder whereby generalized hair shedding
(teloptosis) of greater than 20% existed than observed in FPHL [84].
Although combination occurrences of FPHL and CTE represent the
most common manifestations of female hair loss, other presentations,
such as cycle delays and lag phases [85], complicate diagnoses and
treatments as they may possess slightly different variant mechanisms
of hair loss.
Recent global gene expression studies [86-88] reported that a
defect in conversion of stem cells to daughter progenitor stem cells in
the bulge area might play a role in the pathogenesis of male AGA. In
haired male humans [88], isolated bulge and their progeny secondary
stem cells possessed high levels of preferential expression of the
intermediate filament protein KRT [15] and cell surface markers
CD200+, CD34+ and ITGA6+. At the end of normal cycling during catagen, secondary germ cells arose as progeny from bulge stem cells,
expressed similar KRT15 filament proteins and CD 200+, CD34+
and ITAG6+ bulge cell markers, and were immediately responsible
for the formation of the new lower hair follicle at early anagen. In
bald male humans, however, isolated cell populations of secondary
germ progenitor cells were greatly diminished, possibly due to lack of
replenishment from bulge cells, and potentially resulted in reduced
populations of matrix stem cells which were responsible for hair
shaft production. The roles of these genomic mechanisms in the
pathogenesis of male AGA have yet to be identified in FPHL and CTE
conditions [89].
Evaluation of Hair Loss
The human scalp is predetermined to have approximately 100,000
terminal hair follicles at birth. At any time, most follicles are either
in the growing anagen phase (90-95%), regressive catagen phase (<
1%), or quiescent telogen phase (≃10%). At the end of telogen, about
100 miniaturized shafts are shed and the same number entered the
anagen cycle.
The diagnosis of male AGA, female FPHL or CTE is based
primarily on the basis of medical history, physical examination,
context of age and occupation, and clinical importance with the need
for biopsies rarely needed. Because of their less common occurrence,
the reader is referred to other reviews [90] for further discussion of
diffuse alopecia areata, an autoimmune disease, cicatricial traumatic
alopecia and other less infrequent diseases.
Non-Surgical Treatment Options
Although hair transplantation or surgical reduction procedures,
and hairpieces are viable options for some patients, a number of nonapproved
USA Food and Drug Administration (FDA) treatments for
AGA has been available for AGA that included topical formulations,
oral supplements, and prescription medications and low-level light
[92-95]. Although their individual mechanisms of action have not
been fully elucidated with few clinical controlled trials [96-98], they
are offered as monotherapy or as combination therapies. Two FDAapproved
drugs have been available, however, for the treatment
of male and female pattern hair loss but differ in mechanisms of
action and route of administration. They are prescribed as standalone
treatment options to maintain as much coverage as possible
or combined with hair transplantation surgery to reduce telogen
shedding. Although both drugs retard hair loss and stimulate anagen
growth, neither restores hair loss completely.
Minoxidal, a piperidinopyrmidine derivative, was reported
to produce unexpectedly hypertrichosis in 24%-100% of patients
who were being treated for refractory cases of hypertension as an
oral medication in the 1970s [99-101]. Subsequent dose-dependent
placebo controlled studies determined that the FDA-approved
marketing of topical 2% or 5% concentrated solutions was safe
and effective for temporary AGA improvement in males [102] and
females [103] and for post-hair transplantation [104], appearing to
prolong the duration of anagen, reverse miniaturization, and reduce
postsurgical telogen effluvium. The precise mechanisms of action
may be attributable to the conversion of minoxidal to its active form
of minoxidil sulfate by sulfotransferase enzymes [105], localized
selectively in the lower outer root sheath to its active form(minoxidil
sulfate) [106] that producedarteriolar vasodilation as a potassium
channel opener [107], increased Doppler cutaneous blood flow up
to an hour after application [108], stimulated vascular endothelial
growth factors [109], and promoted dermal papilla cells proliferation
[110]. Although debate continues whether minoxidil prolongs
anagen or shortens telogen, clinical studies in balding men have
shown increased anagen/telogen ratios after 12 months [111], as well
as improved mean hair diameters after 12 months at 4 months [112].
Reversible side effects such as contact dermatitis and unwanted hair
in both men and women have been well documented [113,114].
Finasteride, a member of the azasteroid family, was originally
introduced for the treatment of benign prostatic hypertrophy and was
FDA-approved in 1997 [115] in a 1 mg oral dose for male pattern AGA.
Initial randomized placebo-controlled trials demonstrated prolonged
hair growth to the balding vertex [116] and frontal scalp [117]. The
mechanisms of action have been ascribed to its selective inhibition of
type 2 5α-reductase isoenzyme that converts testosterone to DHT. The
60-70% reduction in DHT [118] is believed to reduce miniaturization
and increase the anagen/telogen ratios [119]. Finasteride was suggested
not only to induce the prolongation of anagen hairs [120], but also the
reduced expression of caspases and apoptosis inhibitors to activate
anagen growth [121,122]. The prolonged use of finasteride resulted
in minimal side effects as compared to placebo except for slight
increase with sexual complaints of less libido and erectile/ejaculatory
dysfunctions [116] and unwanted hair in both sexes [120-123]. The
Prostate Cancer Prevention Trial for patients taking finasteride 5mg/
day reported a relative reduction in the prevalence of prostate cancer
but higher Gleason scores on positive biopsies [124]. Currently, there
is no FDA-approval for its use in the treatment of alopecia in women.
Finasteride is contraindicated in child-bearing aged females, unless
effective birth control measures are used, because of feminization of
a male fetus. Some but not all FPHL-affected women respond to 5
α-reductase inhibitors, indicating the relative minimal significance of
DHT as a primary cause of hair thinning in women [79].
Figure 7
Figure 7
Chest X-ray oblique view for reverse NUSS with moon bridge stabilization.
Controversial Growth Factor Approaches for AGA
With interest in stem cell niches [125] and their growth factors
[126] for cellular proliferation and differentiation, new current
strategies have begun to focus on either platelet-rich plasma (PRP)
[127-130] or adipose tissue containing natural stem cell populations
[55,131,132] for the treatment of female and male AGA. Although
the signaling and transcriptional factors of growth factors have been
studied in nascent and adult phases of the germinal hair cycle, there
have been no recent clinical controlled trials to verify the safety and
efficacy of such protocols.
Platelet-rich plasma contains a high number of platelet particles
that are concentrated above levels normally found in whole blood.
Experimental studies [133,134] have demonstrated that activated
platelets release a number of potent growth factors that include
platelet-derived growth factor (PDGF), fibroblast growth factor
(FGF), hepatocyte growth factor (HGF), transforming growth factor
(TGF), vascular endothelial growth factor (VEGF), epithelial growth
factor (EGF), and insulin-like growth factor (IGF). Growth factors
are believed to be generally responsible for angiogenesis [135], stem
cell proliferation and differentiation [136,137], and anti-apoptotic
properties [138], but specifically for hair follicles, anagen induction
[129], proliferation of dermal papilla cells [139], cyclic growth
[140], and follicle development [141]. Furthermore, Guisti’s in vitro
dose-dependent study [142] indicated the optimal concentration of
activated platelets in PRP to stimulate angiogenesis occurred between
1.5-3.0 x 106 cellular particles/µl.
Although a few published clinical trials without remarkable
side effects have reported 1) improved survival and density of PRPtransplanted
hair follicles in males [127], 2) increased thickness of
PRP-dalteparin/protamine stimulated follicles in thinning hair in
both sexes [128], or 3) increased mean numbers and diameters of
CD34+ PRP stimulated follicles in male and female pattern hair loss
patients [130], further larger, controlled, standardized split scalp
studies will need to be completed to substantiated the safety and
clinical efficacy in both male and female patients with different stages
of hair loss (Figure 3).
The use of autologous fat grafting with or without enhancement
with growth factors from PRP or stromal vascular fractions (SVF)
has been anecdotally reported (Figures 4, 5). At this time, there are
no on-going IRB-approved trials with standardized protocols for
harvesting, processing and delivery of adipose tissue, PRP or SVF,
including optimal dosing and treatment ratios. Currently there exist
no FDA-approved Adipose Stem Cells isolation devices, procedures,
or clinics in the United States.
Figure 3
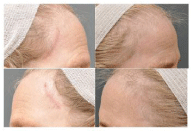
Figure 3
A) This 63 year old Caucasian female presented with telogen
effluvium after a lateral brow lift through post-trichial incisions 1 cm behind
the anterior temporal hair lines. The patient was not on any hormonal
replacement therapy and has never been on Minoxidal treatments. The
exposed scars were visible and of cosmetic concern. B) The patient
underwent one session of micro-needling (three stampings at 1.5 mm, 1.0
mm and 0.5 mm depths) to the entire fronto-temporal hairline and extending
about 5cm into the hair-bearing scalp and simultaneous injection of 4 ml
PRP (0.1 ml/1 cm2
; 40-50 marked treatment squares, 2.4 x 106
platelets/
µl; Harvest-Terumo Technologies Corp., Plymouth, MA). Xylocaine 0.5%
with epinephrine (25 ml) was injected in the subdermal space as the local
anesthesia prior to the procedure. New vellus and terminal hair growth were
observed anterior to the surgical sca
Figure 4
Figure 4
This 64 year old African-American female presented with chronic
telogen effluvium to the entire scalp, especially to the vertex. The patient
was on premarin replacement therapy and had never received Minoxidal
treatments. B) The patient underwent one session of micro-needling (three
stampings at 1.5 mm, 2.0 mm and 2.5 mm depths to the entire fronto-temporoparietal,
and vertex scalp). Immediately thereafter, 30 ml of fat, harvested at
the hip rolls by LipoGems system (Lipogems International S.R.L., Milan IT)
was combined with 3 ml of PRP ( (1.2 x 106 platelets/µl, Harvest-Terumo
Technologies Corp., Plymouth, MA),and injected in the sub-aponeurotic
plane (0.2 ml/1 cm2
) under 150 marked treatment squares. Xylocaine 0.5%
with epinephrine (35 ml) was injected in the sub-aponeurotic space prior to
the procedure. At six months, new vellus and terminal hair was observed
throughout the treated scalp with an increase hair count by computerized
microscopy (Canfield Imaging, Parsippany, NJ).
Figure 5

Figure 5
A) This 72 year old Caucasian female presented with chronic
telogen effluvium to the entire scalp since her mid-twenties. The patient was
not on hormone replacement therapy and had never received Minoxidal
therapy. B) The patient underwent one session of fat grafting combined
with stromal vascular fraction (SVF) to the thinning scalp. A 3-4 mm blunt
tip Mercedes cannula collected about 300 ml of lipoaspirate in 60 ml
Toomey syringes. Five ml of SVF was obtained by enzymatic separation
(Cytori Therapeutics, Inc., San Diego CA). An aliquot of the lipoaspirated
was processed in the PureGraft Filter system (PureGraft, San Diego, CA)
to obtain about 30 ml of purified fat. The PureGraft processed fat was
combined with the 5 ml SVF and injected in 0.2 ml aliquots/1 cm2 square
and injected in the sub-aponeurotic space under approximately 100 marked
treated squares. Xylocaine 0.5% with epinephrine (35-50 ml) was injected in
the sub-aponeurotic space prior to the procedure. New vellus and terminal
hairs were observed 1 year after treatment with an increased hair count by
computerized microscopy (Canfield Imaging, Parsippany, NJ).
Conclusion
The goals of the present chapter was to provide an overview of the importance of stem cell niches in both epidermal and mesenchymal lineages that give rise to specific structures within the nascent mammalian hair follicles and subsequent post-natal follicles. This orchestrated design of recapitulation through triggering of molecular cross-talking signals and transcription factors regulates the various defined cycling stages from miniaturized to fully-developed hair follicles. As differential expressions of stem-cell-associated markers are identified and more functionally understood, an accumulated body of information will provide researchers and clinicians an improved understanding of normal cycling, androgenetic hair loss and more uncommon entities as autoimmune types of alopecia. It is anticipated that evidence-base treatments will continue to be developed with safe and reliable drugs, growth factors and possibly, in the near future, FDA-approved approaches utilizing PRP and stem cells to address hair loss and emotional stress for millions of men and women.
References
- Schofield R. The relationship between the spleen colony-forming cell and the haemopoietic stem cell. Blood Cells. 1978; 4: 7-25.
- Morrison SJ, Kimble J. Asymmetric and symmetric stem cell divisions in development and cancer. Nature. 2006; 441: 1068-1074.
- Fuchs E. The Tortoise and the Hair: Slow Cycling Cells in the Stem Cell Race. Cell. 2009; 137: 811-819.
- Blanpain C, Fuchs E. Epidermal stem cells of the skin. Annu Rev Cell Dev Biol. 2006; 22: 339-373.
- Voog J, Jones D. Stem cells and the niche: a dynamic duo. Cell Stem Cell. 2010; 6: 103-115.
- Paus R, Rover MS, Van Der Veen C. A comprehensive guide for the recognition and classification of distinct stages of hair follicle morphogenesis. J Invest Dermatol. 1999; 113: 523-532.
- Paus R, Cotsarelis G. The biology of hair follicles. N Engl J Med. 1999; 341: 491-497.
- Nowak JA, Polak L, Pasolli HA. Hair follicle stem cells are specified and function in early skin morphogenesis. Cell Stem Cell. 2008; 3: 33-43.
- Schmidt-Ullrich R, Paus R. Molecular principles of hair follicle induction and morphogenesis. BioEssays. 2005; 27: 247-261.
- Cotsarelis G. Epithelial stem cells: folliculocentric view. J Invest Dermatol. 2006; 126: 1459-1468.
- Millar SE. Molecular mechanisms regulating hair follicle development. J Invest Dermatol. 2002; 118: 216-225.
- Tumbar T, Guasch G, Greco V. Defining the epithelial stem cell niche in skin. Science. 2004; 303: 359-363.
- Hardy MH. The secret lift of the hair follicle. Trends Genet. 1992; 8: 55-61.
- Reddy S, Andl T, Bagasra A. Characterization of Wnt gene expression in developing and postnatalhair follicles and identification of Wnt5a as a target of Sonic hedgehog in hair follicle morphogenesis. Mech Dev. 2001; 107: 69-82.
- Andl T, Reddy ST, Gaddapara T. Wnt signals are required for the initiation hair follicle development. Dev Cell. 2002; 2: 643-653.
- Schmidt-Ullrich R, Aebischer T. Requirement of NF-kappaβ/Rel for the development of hair follicles and other epidermal appendices. Development. 2001; 128: 3843-3853.
- Jamora C, DasGupta R, Kocieniewski P. Links between signal transduction, transcription and adhesion in epithelial bud development. Nature. 2003; 422: 317-322.
- Mill P, Mo R, Fu H. Sonic hedgehog-dependent activation of Gli2 is essential for embryonic hair follicle development. Genes Dev. 2003; 17: 282-294.
- Yang CC, Cotsarelis G. Review of hair follicle dermal cells. J Dermatol Sci. 2010; 57: 1-10.
- McMahon AP, Ingham PW, Tabinj CJ. Developmental roles ad clinical significance of hedgehog signaling. Curr Top Dev Biol. 2003; 53: 1-114.
- Vidal VP, Chaboissier MC, Lutzkendorf S. Sox9 is essential for outer root sheath differentiation and the formation of the hair stem cell compartment. Curr Biol. 2005; 15: 1340-1351.
- Rhee H, Polak L, Fuchs E. Lhx2 maintains stem cell character in hair follicles. Science. 2006; 312: 1946-1949.
- Cotsarelis G, Sun TT, Lavcker RM. Label-retaining cells reside in the bulge area of the pilosebaceous unit: implications for follicular stem cells, hair cycle, and skin carcinogenesis. Cell. 1990; 61: 1329-1337.
- Zhang YV, Cheong J, Ciapurin N. Distinct self-renewal and differentiation phases in the niche of infrequently dividing hair follicle stem cells. Cell Stem Cell. 2009; 5: 267-278.
- Rendl M, Lewis L, Fuchs E. Molecular dissection of mesenchymal-epithelial interactions in the hair follicle. PLoS Biol. 2005; 3: 1910-1924.
- Blanpain C, Lowry WE, Geoghegan A. Self-renewal, multipotency, and the existence of two cell populations within an epithelial stem cell niche. Cell. 2004; 118: 635-648.
- Liu Y, Lyle S, Yang Z. Keratin 15 promoter targets putative epithelial stem cells in the hair follicle bulge. J Invest Dermatol. 2003; 121: 963-968.
- Ellis T, Gambardella L, Horcher M. The transcriptional repressor CDP (Cultl1) is essential for epithelial cell differentiation of the lung and the hair follicle. Genes Dev. 2001; 15: 2307-2319.
- Kaufman CK, Zhou P, Pasolli HA. GATA-3: an unexpected regulator of cell lineage determination in skin. Genes Dev. 2003; 17: 2108-2122.
- Peters EM, Botchkarev VA, Muller-Rover S. Developmental timing of hair follicle and dorsal skin innervation in mice. J Comp Neurol. 2002; 448: 28-52.
- Philpott MP, Paus R. Principles of hair follicle morphogenesis. In: Chuong CM, editor. Molecular Basis of Epithelial Appendage Morphogensis. Austin, TX: R.G. Landes. 1998; 75.
- Petiot A, Conti FJ, Grose R. A crucial role for Fgfr2-III signalling in epidermal development and hair follicle patterning. Development. 2003; 130: 5493-5501.
- Botchkarev VA, Botchkareva NV, Roth W. Noggin is a mesenchymally derived stimulator of hair-follicle induction. Nat Cell Biol. 1999; 1: 158-164.
- Chuong CM, Cotsarelis G, Stenn K. Defining hair follicles in the age of stem cell bioengineering. J Invest Dermatol. 2007; 127: 2098-2100.
- St-Jacques B, Dassule HR, Karavanova I. Sonic hedgehog signaling is essential for hair development. Curr Biol. 1998; 8: 1058-1068.
- Karisson L, Bondjers C, Betsholtz C. Roles for PDGF-A and sonic hedgehog in development of mesenchymal components of the hair follicle. Development. 1999; 126: 2611-2621.
- Ito M, Sato Y. Dynamic ultrastructural changes of the connective tissue sheath of human hair follicles during hair cycle. Arch Dermatol Res. 1990; 282: 434-441.
- Tobin DJ, Gunin A. Magerl M. Plasticity and cytokinetic dynamics of the hair follicle mesenchyme: implications for hair growth control. J Invest Dermal. 2003; 120: 895-904.
- McElwee KJ, Kissling S, Wenzel E. Cultured peribulbar dermal sheath cells can induce hair follicle development and contribute to the dermal sheath and dermal papilla. J Invest Dermatol. 2003; 121: 1267-1275.
- Feutz AC, Barrandon Y, Monard D. Control of thrombin signaling through P13K is a mechanism underlying plasticity between hair follicle dermal sheath and papilla cells. J Cel Sci. 2008; 121: 1435-1443.
- Greco V, Chen T, Rendl M. A two-step mechanism for stem cell activation during hair regeneration. Cell Stem Cell. 2009; 4: 155-169.
- Ito M, Kizawa K, Hamada K. Hair follicle stem cells in the lower bulge form the secondary germ, a biochemically distinct but functionally equivalent progenitor cell population, at the termination of catagen. Differentiation 2004; 72: 548-557.
- Zhang Y, Andl T, Yang S. Activation of {beta}-catenin signaling programs embryonic epidermis to hair follicle fate. Development. 2008; 135: 2161-2172.
- Horsley V, Aliprantis AO, Polak L. NFAYcl balances quiescence and proliferation of skin stem cells. Cell. 2008; 132: 299-310.
- Kobielak K, Pasolli HA, Alonso L. Defining BMP functions in the hair follicle by conditional ablation of BMP receptor IA. Proc Natl Acad Sci USA. 2007; 104: 10063.
- Lowry WE, Blanpain C, Nowak JA. Defining the impact of beta-catenin/Tef transactivation on epithelial stem cells. Genes Dev. 2005; 19: 1596-1611.
- Waghmare SK, Bansal R, Lee J. Quantitative proliferation dynamics and random chromosome segregation of hair follicle stem cells. EMBO J. 2008; 27: 1309-1320.
- Panteleyev AA, Jahoda CA, Christiano AM. Hair follicle predetermination. J Cell Sci. 2001; 114: 3419-3431.
- Sbarti A, Accorsi D, Berati D. Subcutaneous adipose tissue classification. Eur J Histochem. 2010; 54: e48.
- Schmidt B, Horsley V. Unraveling hair follicle-adipocyte communications. Expert Dermatol. 2012; 21: 827-830.
- Hansen LS, Coggle JE, Wells J. The influence of the hair cycle on the thickness of mouse skin. Anat Rec. 1984; 210: 569-573.
- Hausman GJ, Campion DR, Richardson RL. Adipocyte development in the rat hypodermis. Am J Anat. 1982; 161: 85-100.
- Rodeheffer MS, Birsoy K, Friedman JM. Identification of white adipocyte progenitor cells in vivo. Cell. 2008; 135: 240-249.
- Rosen ED, Spiegelman BM. Molecular regulation of adipogenesis. Annu Rev Cell Dev Biol. 2000; 16: 145-171.
- Festa E, Fretz J, Berry R. Adipocyte lineage cells contribute to the skin stem cell niche to drive hair cycling. Cell. 2011; 146: 761-771.
- Hesslein DGT, Fretz JA, Xi Y. Ebfl-dependent control of the osteoblast and adipocyte lineages. Bone. 2009; 44: 537-546.
- Sasaki GH. Plastic surgery update on the biology of fat cells and adipose-derived stem cells for fat grafting. Open Access Library Journal. 2015; 2: 1-26.
- Plikus M, Mayer J, de la Cruz D. Cyclic dermal BMP signalling regulates stem cell activation during hair regeneration. Nature. 2008; 451: 340-344.
- Zhang J, He X-C, Tong WG. Bone morphogenetic protein signaling inhibits hair follicle anagen induction by restricting epithelia stem/progenitor cell activation and expansion. Stem Cells. 2006; 24: 2826-2839.
- Hamilton JB. Patterned loss of hair in man: Types and incidence. AnnNY Acad Sci. 1951; 53: 708-728.
- Olsen EA. Female pattern hair loss. In: Blum-Peytavi U, Tosti A, Whiting D, editors. Hair Growth and Disorders. Berlin Heidelberg: Springer. 2008: 171-186.
- Cash TF. The psychological effects of androgenetic alopecia in men. J Am Acad Dermatol. 1992; 26: 929-931.
- Cash TF, Price VH, Savin RC. Psychological effects of androgenetic alopecia on women: Comparisons with balding men and with female control subjects. J Am Acad Dermatol. 1993; 29: 568-575.
- Ellis JA, Stebbing M, Harrap SB. Genetic analysis of male pattern baldness and 5α-reductase genes. J Invest Dermatol. 1998; 110: 849-853.
- Griffen JE, Wilson JD. The androgen resistance syndromes: 5α-reductase deficiency, testicular feminization, and related disorders. In: Scriver CR, Beaudet AL, Sly WS, Valle D, editors. The Metabolic Basis of Inherited Disease. 6th ed. New York, NY. McGraw-Hill. 1989; 2: 1919-1944.
- Feinstein R. Androgenetic alopecia. 2015.
- Sawaya ME, Price VH. Different levels of 5alpha-reductase type I and II, aromatase, and androgen receptor in hair follicles of women and men with androgenetic alopecia. J Invest Dermatol. 1997; 109: 296-300.
- Kaufman KD. Androgens and alopecia. Mol Cell Endocrinol. 2002; 198: 89-95.
- Alsantali A, Shapiro J. Androgens and hair loss. Curr Opin Endocrinol Diabetes Obes. 2009; 16: 246-253.
- Martinon F, Mayor A, Tschopp J. The inflammasomes: guardians of the body. Annu Rev Immunol. 2009; 27: 229-265.
- de Rovero Vaccaro J, Swaya ME, Brand F. Caspase-1 level is higher in the scalp in androgenetic alopecia. Dermatol Surg. 2012; 38: 1033-1039.
- Rattew JA, Huet-Hudson YM, Marriott I. Testosterone reduces macrophage expression in the mouse of toll-like receptor 4, a trigger for inflammation and innate immunity. Bio Reprod. 2008; 78: 432-437.
- Bergsbaken T, Fink SL, den Hartigh AB. Coordinated host responses during pyroptosis: caspase-1-dependent lysosome exocytosis and inflammatory cytokine maturation. J Immunol. 2011; 871: 2748-2754.
- Norwood O. Male pattern baldness: Classification and incidence. South Med J. 1975; 68: 1359-1365.
- Leavitt M. Understanding and management of female pattern alopecia. Facial Plast Surg. 2008; 24: 414-417.
- Yip L, Rufaut N, Sinclair R. Role of genetics and sex steroid hormones in male adrogenetic alopecia and female pattern hair loss: an update of what we now know. Australas J Dermatol. 2011; 52: 81-881.
- Hernadez BA. Is androgenetic alopecia a result of endocrine effects on the vasculature? Med Hypotheses. 2004; 62: 438-441.
- Orme S, Cullen DR, Messenger AG. Diffuse female hair loss: are androgens necessary? Br J Dermatol. 1999; 141: 521-523.
- Price VH, Roberts JL, Hordkinsky M. Lack of efficacy of finasteride in postmenopausal women with androgenetic alopecia. J Am Acad Dermatol. 2000; 43: 768-776.
- Gordon KA, Tosti A. Alopecia: evaluation and treatment. Clin Cosmet Investig Dermatol. 2011; 4: 101-106.
- Messenger AG, Sinclair R. Follicular miniaturization in female pattern hair loss: clinicopathological correlations. Br J Dermatol. 2006; 155: 926-930.
- Blume-Peytavi U, Blumeyer A, Tosti A. S1 guideline for diagnostic evaluation in adrogenetic alopecia in men, women, and adolescents. Br J Dermaol. 2011; 164: 5-15.
- Whiting DA. Chronic telogen effluvium: increased scalp hair shedding in middle-aged women. J Am Acad Dermatol. 1996; 35: 899-906.
- Trueb RM. Das idiopathische chronische telogen effluvium der frau. Hautarzt. 2000; 51: 899-905.
- Rebora A. Pathogenesis of androgenetic alopecia. J Am Acad Dermatol. 2004; 50: 777-779.
- Inoue K, Aoi N, Sato T. Differential expression of stem-cell-associated markers in human hair follicle epithelial cells. Lab Invest. 2009; 89: 844-856.
- Rittie L, Stroll SW, Kang S. Hedgehog signaling maintains hair follicle stem cell phenotype in young and aged human skin. Aging Cell. 2009; 8: 738-751.
- Garza LA, Yang C-C, Zhao T. Bald scalp in men with androgenetic alopecia retains hair follicle stem cells but lacks CD200-rich and CD34-positive hair follicle progenitor cells. J Clin Invest. 2011; 121: 613-622.
- Ohyama M, Terunuma A, Tock CL. Characterization and isolation of stem cell-enriched human hair follicle bulge cells. J Clin Invest. 2006; 116: 249-260.
- Olsen EA, Messenger AF, Shapiro J. Evaluation and treatment of male and female pattern hair loss. J Am Adad Dermatol. 2005; 52: 301-311.
- Rogers NE, Avram MR. Medical treatments for male and female pattern hair loss. J Am Acad Dermatol. 2008; 59: 547-566.
- Lengg N, Heidecker B, Seifert B. Dietary supplement increases anagen hair rate in women with telogen effluvium: results of a double-blind, placebo-controlled trial. Therapy. 2007; 4: 59-65.
- Glynis A. A double-blind, placebo-controlled study evaluating the efficacy of an oral supplement in women with self-perceived thinning hair. J Clin Aesthet Dermatol. 2012; 5: 28-34.
- Hornfeldt CS, Holland M. The safety and efficacy of a sustainable marine extract for the treatment of thinning hair: A summary of new clinical research and results from a panel discussion on the problems of thinning hair and current treatments. J Drug Derm. 2015; 14: 15-22.
- Avram MR, Leonard RT, Jr, Epstein ES. The current role of laser/light sources in the treatment of male and female pattern hair loss. J Cosmet Laser Ther. 2007; 9: 27-28.
- van Zuuren EJ, Fedorowicz A, Carter B. Evidence-based treatments for female pattern hair loss: A summary of a Cochrane systematic review. Br J Dermatol. 2012; 167: 995-1010.
- van Zuuren EJ, Fedorowicz A, Carter B. Interventions for female pattern hair loss. Cochraine Database Syst Rev. 2012; 5: CD007628.
- Irwig MS. Safety concerns regarding 5α reductase inhibitors for the treatment of androgenetic alopecia. Curr Opin Endocrinol Diabetes Obes. 2015; 22: 248-0253.
- Jacomb RG, Brunnberg FJ. The use of minoxidil in the treatments of severe essential hypertension: a report on 100 patients. Clin Sci Mol Med Suppl. 1976; 3: 579s-581.
- Dargie HJ, Dollery CT, Daniel J. Minoxidil in resistant hypertension. Lancet. 1077; 2: 515-518.
- Burton JL, Marshall A. Hypertrichosis due to minoxidil. Br J Dermatol. 1979; 101: 593-595.
- Olsen EA, Dunlap FE, Funicella T. A randomized clinical trial of 5%topical minoxidil versus 2% topical minoxidil and placebo in the treatment of adrogenetic alopecia in man. J Am Acad Dermatol. 2002; 47: 377-385.
- Lucky AW, Picquadio DJ, Ditre CM. A randomized, placebo-controlled trial of 5% and 2% topical minoxidil solutions in thetreatment of female pattern hair loss. J Am Acad Dermatol. 2004; 50: 541-543.
- Avram MR, Cole JP, Gandelman M. The potential role of minoxidil in the hair transplantation setting: roundtable consensus meeting of the9th annual meeting of the International Society of Hair RestorationSurgery. Dermatol Surg. 2002; 28: 894-900.
- Buhl AE, Waldon DJ, Baker CA. Minoxidil sulfate is the activemetabolite that stimulates hair follicles. J Invest Dermatol.1990; 95: 553-557.
- Dooley TP, Walker CJ, Dietz AJ. Localization of minoxidilsulfotransferase in rat liver and the outer root sheath of anagen pelageand vibrissa follicles. J Invest Dermatol. 1991; 96: 65-70.
- Meisheri KD, Cipkus LA, Taylor CJ. Mechanism of action of minoxidil sulfate-induced vasodilation: a role for increased K+ permeability. J Pharmocol Exp Ther. 1998; 245: 751-760.
- Wester RC, Maibach HI, Guy RH. Minoxidil stimulates cutaneousblood flow in human balding scalp: pharmacodynamics measured bylaser Doppler velocimetry and photopulse plethysmography. J InvestDermatol. 1984; 82: 515-517.
- Lachgar S, Chareron M, Gail Y. Minoxidil upregulates theexpression of vascular endothelial growth factor in human dermalpapilla cells. Br J Dermatol. 1998; 138: 407-411.
- Han JH, Kwon OS, Chung JH. Effect of minoxidil on proliferation and apoptosis in dermal papilla cells of human follicle. J Dermatol Sci. 2004; 34: 91-98.
- Abell E. Histologic response to topically applied minoxidil in male-patterned alopecia. Clin Dermatol. 1988; 6: 191-194.
- Headington JT, Novak E. Clinical and histologic studies of male pattern baldness treated with topical minoxidil. Curr Ther Res. 1984; 36: 1098-1106.
- Rietschel RL, Duncan SH. Safety and efficacy of topicalminoxidil in themanagement of androgenetic alopecia. J Am Acad Dermatol. 1987; 16: 677-685.
- Dawber RP, Rundegren J. Hypertrichosis in females applying minoxidil topical solution and in normal controls. J Eur Acad Dermatol. 19887; 16: 677-685.
- Roberts JL, Fiedler V, Imperato-McGinley J. Clinical dose ranging studies with finasteride, a type2 5-alpha reductase inhibitor, in men with male pattern hair loss. J Am Acad Dermatol. 1999; 41: 555-563.
- Finasteride Male Pattern Hair Loss Study Group. Long-term (5 year) multinational experience with finasteride 1 mg in the treatment of men with androgenetic alopecia. Eur J Dermatol. 2002; 12: 38-49.
- Leyden J, Dunlap F, Miller B. Finasteride in treatment of men with frontal male pattern hair loss. J Am Acad Dermatol. 1999; 40: 930-937.
- Drake L, Hordinsky M, Fiedler V. The effects of finasteride on scalp skin and serum androgen levels in men with androgenetic alopecia. J Am Acad Dermatol. 1999; 41: 550-554.
- Whiting DA, Waldstreicher HA, Sanchez M. Measuring reversal of hair miniaturization in androgenetic alopecia by follicular counts in horizontal sections of serial scalp biopsies: results of finasteride 1 mg treatment of men and postmenausal women. J Investig Dermatol Symp Proc. 1999; 4: 282-284.
- Tosti A, Piraccini BM. Finasteride and the hair cycle. J Am Acad Dermatol. 2000; 42: 848-849.
- Sawaya Me, Blume-peytavi U, Mullins DL. Effects of finasteride on apoptosis and regulation of human hair cycle. J Cutan Med Surg. 2002; 6: 1-9.
- de Rivero Vaccari JP, Sawaya ME. Caspase-1 level is higher in the scalp in androgenetic alopecia. Dermatol Surg. 2012; 38: 1033-1039.
- Libecco JF, Bergfelde WF. Finasteride in the treatment of alopecia. Expert Opin Pharmacother. 2004; 5: 933-940.
- Thompson IM, Goodman PJ, Tangen CM. The influence of finasteride on the development of prostate cancer. N Engl J Med. 2003; 349: 215-224.
- Goldstein J, Valerie H. Home sweet home: Skin stem cell niches. Cell Mol Life Sci. 2012; 69: 2573-2582.
- Kilroy GE, Foster S, Wu X. Cytokine profile of human adipose-derived stem cells: Expression of angiogenic, hematopoietic, and pro-inflammatory factors. J of Cell Physiol. 2007; 212: 702-709.
- Uebel CO, da Silva JB, Cantarelli D. The role of platelet plasma growth factors in male pattern baldness surgery. Plast Reconstr Surg. 2006; 118: 1458-1466.
- Takikawa M, Nakamura S, Nakmura S. Enhanced effect of platelet-rich plasma containing a new carrier on hair growth. Dermatol Surg. 2011; 37: 1721-1729.
- Li ZI, Choi HI, Choi DK. Autologous platelet-rich plasma: A potential therapeutic tool for promoting hair growth. Dermatol Surg. 2012; 38: 1040-1046.
- Kang JS, Zheng Z, Choi MJ. The effect of CD34+ cell-containing autologous platelet-rich plasma injection on pattern hair loss: A preliminary study. J Eur Dermatol Venereol. 2014; 28: 72-79.
- Lee GR, Sasaki GH. PRP for hair loss. Med Esthetics. 2014: 28-32.
- Sasaki GH. Micro-needling: Induced collagen formation and delivery system for skin and hair enhancement. Teaching Course 414. Annual Meeting of the American Society for Aesthetic Plastic Surgery. 2016.
- Eppley BL, Petrzak WS, Blanton M. Platelet-rich plasm: a review of biology and applications in plastic surgery. Plast Reconstr Surg. 2004; 114: 1502-1508.
- Weibrich G, Kleis SWK, Hitzler WE. Growth factor levels in platelet-rich plasma and correlations with donor age, sex, and platelet count. J Craniomaxillofac Surg. 2002; 30: 97-102.
- Bir SC, Esaki J, Marui A. Angiogenic properties of sustained release platelet-rich plasma: characterization in vitro and in the ischemic hind limb of the mouse. J Vasc Surg. 2009; 50: 870-8798:e2.
- Kakudo N, Minakata T, Mitsui T. Proliferation-promoting effect of platelet-rich plasma on human adipose-derived stem cells and human dermal fibroblasts. Plast Reconstr Surg. 2008; 122: 1352-1360.
- Cervelli V, Scioli MG, Gentile P. Platelet-rich plasma greatly potentiates insulin-induced adipogenic differentiation of human adipose-derived stem cells through a sere/threonine kinase Akt-dependent mechanism and promotes clinical fat graft maintenance. Stem Cells Transl Med. 2012; 1: 206-220.
- Fukaya Y, Kuroda M, Aoyagi Y. Platelet-rich plasma inhibits the apoptosis of highly adipogenic homogeneous pre-adipocytes in an in vitro culture system. Exp Mol Med. 2012; 44: 330-339.
- Itami S, Kurata S, Takayasu S. Androgen induction of follicular epithelial cell growth is mediated via insulin-like growth factor-1 from dermal papilla cells. Biochem Biophys Res Commun. 1995; 21: 988-995.
- Jindo T. Tsuboi R, Takamori K. Local injection of hepatocyte growth factor/scatter factor (HGF/SF) alters cyclic growth of murine hair follicles. J Invest Dermatol. 1998; 110: 338-342.
- Guo L, Degenstein L, Fuchs E. Keratinocyte growth factor is required for hair development but not for wound healing. Genes Dev. 1996; 10: 165-175.
- Giusti I, Rughetti A, D’Ascenszo S. Identification of an optimal concentration of platelet gel for promoting angiogenesis in human endothelial cells. Transfusion. 2009; 49: 771-778.